The Flamingo’s Smile (17 page)
Read The Flamingo’s Smile Online
Authors: Stephen Jay Gould

As an example of how this pattern illuminates the larger question, we have used our distinction of mottled and ribby to prove, for the first time, that the unconventional hypothesis advanced by most
Cerion
experts is indeed valid: the complex suite of characters defining such basic forms as mottled and ribby can evolve independently many times. We find the same distinction of mottled and ribby on both Little and Great Bahama banks. Conventional wisdom would hold that the mottled snails of both banks represent one stock, while the ribby snails of both banks form a different genealogical group. But Daniel Chung, a student of Woodruff’s, and Simon Tillier, a leading anatomist of land snails at the Paris Museum, have studied the genital anatomy of these snails for us, and have made the following surprising discovery:
both
mottled and ribby snails of Little Bahama Bank share the
same
anatomy, while both mottled and ribby on Great Bahama Bank share a distinctly different set of genital structures. (Genital anatomy is the standard tool for establishing genealogical affinities among land snails. The differences are sufficiently profound and complex to indicate that shared anatomy reflects common descent while shared shell morphology must evolve independently.) Thus, the complex of traits defining mottled and ribby can evolve again and again. We would not have been able to reach this conclusion had we not extracted the pattern of mottled and ribby from a previous chaos of names.
At this point, I think we begin to peer through a glass darkly at the deeper mystery of form. We have shown that a complex set of independent traits can evolve in virtually the same way more than once. I do not see how this can happen if each trait must develop separately, following its own genetic pathway, each time. The traits must somehow be coordinated in
Cerion
’s genetic program; they must be evoked, or “called forth,” together. Some genetic releaser must coordinate the joint appearance of these characters. Does the master genetic program of all
Cerions
encode alternate pathways representing the several basic forms that evolve again and again? The homeotic mutations of insects (see essay 15 in
Hen’s Teeth and Horse’s Toes
) indicate that some such hierarchical system must regulate development, for the production of well-formed parts in the wrong places (legs where antennae should be, for example) indicates that some master switch must regulate all the genes that produce legs, and that higher controls can turn the master switch on in the wrong place or at the wrong time. Likewise, some master switch within
Cerion
’s program might evoke any one of its basic developmental pathways and evolve the set of traits that marks its fundamental forms again and again.
In this way,
Cerion
provides insight into what may be the most difficult and important problem in evolutionary theory: How can new and complex forms (not merely single features of obvious adaptive benefit) arise if each requires thousands of separate changes, and if intermediate stages make little sense as functioning organisms? If genetic and developmental programs are organized hierarchically, as homeotic mutations and the multiple evolution of basic forms in
Cerion
suggest, then new designs need not arise piecemeal (with all the intractable problems attached to such a view), but in a coordinated way by manipulating the master switches (or “regulators”) of developmental programs. Yet so deep is our present ignorance about the nature of development and embryology that we must look at final products (an adult
Drosophila
with a leg where its antenna should be or mottled
Cerions
evolved several times) to make uncertain inferences about underlying mechanisms.
I chose
Cerion
because I thought it might illuminate these large and woolly issues. Yet, although they lurk always in the back of my mind, they are not the source of my daily joy. Little predictions affirmed or small guesses proved wrong and exchanged for more interesting ideas are the food of continual satisfaction.
Cerion
, or any good field project, provides unending stimulation, so long as little puzzles remain as intensely absorbing, fascinating, and frustrating as big questions. Fieldwork is not like the one-hundred-thousandth essay on Shakespeare’s sonnets; it always presents something truly new, not a gloss on previous commentaries.
I remember when we found the first population of living
Cerion agassizi
in central Eleuthera. Our hypothesis of
Cerion
’s general pattern required that two predictions be affirmed (or else we were in trouble): this population must disappear by hybridization with mottled shells toward bank-interior coasts and with ribby snails toward the bank-edge. We hiked west toward the bank-interior and easily found hybrids right on the verge of the airport road. We then moved east toward the bank-edge along a disused road with vegetation rising to five feet in the center between the tire paths. We should have found our hybrids, but we did not. The
Cerion agassizi
simply stopped about two hundred yards north of our first ribby
Cerion
. Then we realized that a pond lay just to our east and that ribby forms, with their coastal preferences, might not favor the western side of the pond. We forded the pond and found a classic hybrid zone between
Cerion agassizi
and ribby
Cerions
. (Ribby
Cerion
had just managed to round the south end of the pond, but had not moved sufficiently north along the west side to establish contact with
C. agassizi
populations.) I wanted to shout for joy. Then I thought, “But who can I tell; who cares?” And I answered myself, “I don’t have to tell anyone. We have just seen and understood something that no one has ever seen and understood before. What more does a man need?”
An eminent colleague, a fine theoretician who has paid his dues in the field, once said to me, only partly facetiously, that fieldwork is one hell of a way to get information. All that time, effort, and money, often for comparatively little when measured against the hours invested. True enough, especially when I count the hours spent drinking Cuban coffee, the one pleasure of my least favorite place, Miami airport. But all the frustration and dull, repetitive effort vanish to insignificance before the unalloyed joy of finding something new—and this pleasure can be savored nearly every day if one loves the little things as well. To say, “We have discovered it; we understand it; we have made some sense and order of nature’s confusion.” Can any reward be greater?
PRETORIA, AUGUST
5, 1984
History’s most famous airplane, Lindbergh’s
Spirit of St. Louis
, hangs from the ceiling of Washington’s Air and Space Museum, imperceptible in its majesty to certain visitors. Several years ago, a delegation of blind men and women met with the museum’s director to discuss problems of limited access. Should we build, he asked, an accurate scale model of Lindbergh’s plane, freely available for touch and examination? Would such a replica solve the problem? The delegation reflected together and gave an answer that moved me deeply for its striking recognition of universal needs. Yes, they said, such a model would be acceptable, but only on one condition—that it be placed directly beneath the invisible original.
Authenticity exerts a strange fascination over us; our world does contain sacred objects and places. Their impact cannot be simply aesthetic, for an ersatz absolutely indistinguishable from the real McCoy evokes no comparable awe. The jolt is direct and emotional—as powerful a feeling as anything I know. Yet the impetus is purely intellectual—a visceral disproof of romantic nonsense that abstract knowledge cannot engender deep emotion.
Last night, I watched the sun set over the South African savanna—the original location and habitat of our australopithecine ancestors. The air became chill; sounds of the night began, the incessant repetition of toad and insect, laced with an occasional and startling mammalian growl; the Southern Cross appeared in the sky, with Jupiter, Mars, and Saturn ranged in a line above the arms of Scorpio. I sensed the awe, fear, and mystery of the night. I am tempted to say (describing emotions, not making any inferences about realities, higher or lower) that I felt close to the origin of religion as a historical phenomenon of the human psyche. I also felt kinship in that moment with our most distant human past—for an
Australopithecus africanus
may once have stood, nearly three million years ago, on the same spot in similar circumstances, juggling (for all I know) that same mixture of awe and fear.
I was then rudely extricated from that sublime, if fleeting, sentiment of unity with all humans past and present. I remembered my immediate location—South Africa, 1984 (during a respite in Kruger Park from a lecture tour on the history of racism). I also understood, in a more direct way than ever before, the particular tragedy of the history of biological views about human races. This history is largely a tale of division—an account of barriers and ranks erected to maintain the power and hegemony of those on top. The greatest irony of all presses upon me: I am a visitor in the nation most committed to myths of inequality—yet the savannas of this land staged an evolutionary story of opposite import.
My visceral perception of brotherhood harmonizes with our best modern biological knowledge. Such union of feeling and fact may be quite rare, for one offers no guide to the other (more romantic twaddle aside). Many people think (or fear) that equality of human races represents a hope of liberal sentimentality probably squashed by the hard realities of history. They are wrong.
This essay can be summarized in a single phrase, a motto if you will:
Human equality is a contingent fact of history
. Equality is not true by definition; it is neither an ethical principle (though equal treatment may be) nor a statement about norms of social action. It just worked out that way. A hundred different and plausible scenarios for human history would have yielded other results (and moral dilemmas of enormous magnitude). They didn’t happen.
The history of Western views on race is a tale of denial—a long series of progressive retreats from initial claims for strict separation and ranking by intrinsic worth toward an admission of the trivial differences revealed by our contingent history. In this essay, I shall discuss just two main stages of retreat for each of two major themes: genealogy, or separation among races as a function of their geological age; and geography, or our place of origin. I shall then summarize the three major arguments from modern biology for the surprisingly small extent of human racial differences.
GENEALOGY, THE FIRST ARGUMENT
Before evolutionary theory redefined the issue irrevocably, early to mid-nineteenth-century anthropology conducted a fierce debate between schools of monogeny and polygeny. Monogenists advocated a common origin for all people in the primeval couple, Adam and Eve (lower races, they then argued, had degenerated further from original perfection). Polygenists held that Adam and Eve were ancestors of white folks only, and that other—and lower—races had been separately created. Either argument could fuel a social doctrine of inequality, but polygeny surely held the edge as a compelling justification for slavery and domination at home and colonialism abroad. “The benevolent mind,” wrote Samuel George Morton (a leading American polygenist) in 1839, “may regret the inaptitude of the Indian for civilization…. The structure of his mind appears to be different from that of the white man…. They are not only averse to the restraints of education, but for the most part are incapable of a continued process of reasoning on abstract subjects.”
GENEALOGY, THE SECOND ARGUMENT
Evolutionary theory required a common origin for human races, but many post-Darwinian anthropologists found a way to preserve the spirit of polygeny. They argued, in a minimal retreat from permanent separation, that the division of our lineage into modern races had occurred so long ago that differences, accumulating slowly through time, have now built unbridgeable chasms. Though once alike in an apish dawn, human races are now separate and unequal.
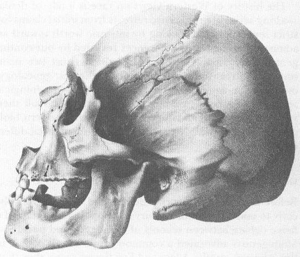
One of John Collins’s excellent lithographs from S.G. Morton’s
Crania Americana
, 1839. Note the subtle “arrangement” chosen (probably unconsciously) to enhance the impression of inferiority in this American Indian skull. The skull is not oriented in the conventional plane, but tilted back, so that the cranium seems lower and less vaulted (therefore smaller and more “primitive”). Compare with next figure of a white male.
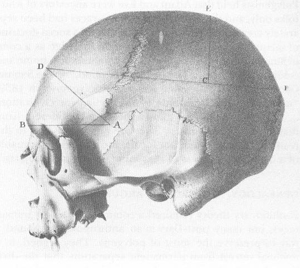
Skull of a white male (with craniometric measures shown) from Morton’s
Crania Americana
. This skull is oriented conventionally (compare with above figure).
We cannot understand much of the history of late nineteenth- and early twentieth-century anthropology, with its plethora of taxonomic names proposed for nearly every scrap of fossil bone, unless we appreciate its obsession with the identification and ranking of races. For many schemes of classification sought to tag the various fossils as ancestors of modern races and to use their relative age and apishness as a criterion for racial superiority. Piltdown, for example, continued to fool generations of professionals partly because it fit so comfortably with ideas of white superiority. After all, this “ancient” man with a brain as big as ours (the product, we now know, of a hoax constructed with a modern cranium) lived in England—an obvious ancestor for whites—while such apish (and genuine) fossils as
Homo erectus
inhabited Java and China as putative sources for Orientals and other peoples of color.
This theory of ancient separation received its last prominent defense in 1962, when Carleton Coon published his
Origin of Races
. Coon divided humanity into five major races—caucasoids, mongoloids, australoids, and, among African blacks, congoids and capoids. He claimed that these five groups had already become distinct subspecies during the reign of our ancestor,
Homo erectus. H. erectus
then evolved toward
H. sapiens
in five parallel streams, each traversing the same path toward increased consciousness. But whites and yellows, who “occupied the most favorable of the earth’s zoological regions,” crossed the threshold to
H. sapiens
first, while dark peoples lagged behind and have paid for their sluggishness ever since. Black inferiority, Coon argues, is nobody’s fault, just an accident of evolution in less challenging environments:
Caucasoids and Mongoloids…did not rise to their present population levels and positions of cultural dominance by accident…. Any other subspecies that had evolved in these regions would probably have been just as successful.
Leading evolutionists throughout the world reacted to Coon’s thesis with incredulity. Could modern races really be identified at the level of
H. erectus?
I shall always be grateful to W.E. Le Gros Clark, England’s greatest anatomist at the time. I was spending an undergraduate year in England, an absolute nobody in a strange land. Yet he spent an afternoon with me, patiently answering my questions about race and evolution. Asked about Coon’s thesis, this splendidly modest man simply replied that he, at least, could not identify a modern race in the bones of an ancient species.
More generally, parallel evolution of such precision in so many lineages seems almost impossible on grounds of mathematical probability alone. Could five separate subspecies undergo such substantial changes and yet remain so similar at the end that all can still interbreed freely, as modern races so plainly do? In the light of these empirical weaknesses and theoretical implausibilities, we must view Coon’s thesis more as the last gasp of a dying tradition than a credible synthesis of available evidence.
GENEALOGY, THE MODERN VIEW
Human races are not separate species (the first argument) or ancient divisions within an evolving plexus (the second argument). They are recent, poorly differentiated subpopulations of our modern species,
Homo sapiens
, separated at most by tens or hundreds of thousands of years, and marked by remarkably small genetic differences.
GEOGRAPHY, THE FIRST ARGUMENT
When Raymond Dart found the first australopithecine in South Africa sixty years ago, scientists throughout the world rejected this oldest ancestor, this loveliest of intermediate forms, because it hailed from the wrong place. Darwin, without a shred of fossil evidence but with a good criterion for inference, had correctly surmised that humans evolved in Africa. Our closest living relatives, he argued, are chimps and gorillas—and both species live only in Africa, the probable home, therefore, of our common ancestor as well.
But few scientists accepted Darwin’s cogent inference because hope, tradition, and racism conspired to locate our ancestral abode on the plains of central Asia. Notions of Aryan supremacy led anthropologists to assume that the vast “challenging” reaches of Asia, not the soporific tropics of Africa, had prompted our ancestors to abandon an apish past and rise toward the roots of Indo-European culture. The diversity of colored people in the world’s tropics could only record the secondary migrations and subsequent degenerations of this original stock. The great Gobi Desert expedition, sponsored by the American Museum of Natural History just a few years before Dart’s discovery, was dispatched primarily to find the ancestry of man in Asia. We remember this expedition for success in discovering dinosaurs and their eggs; we forget that its major quest ended in utter failure because Darwin’s simple inference was correct.
GEOGRAPHY, THE SECOND ARGUMENT
By the 1950s, further anatomical study and the sheer magnitude of continuing discovery forced the general admission that our roots lay with the australopithecines, and that Africa had been our original home. But the subtle hold of unacknowledged prejudice still conspired (with other, more reasonable bases of uncertainty) to deny Africa its continuing role as the cradle of what really matters to us—the origin of human consciousness. In a stance of intermediate retreat, most scientists now argued that Africa had kindled our origin but not our mental emergence. Human ancestors migrated out, again to mother Asia, and there crossed the threshold to consciousness as
Homo erectus
(or so-called Java and Peking man). We emerged from the apes in Africa; we evolved our intelligence in Asia. Carleton Coon wrote in his 1962 book: “If Africa was the cradle of mankind, it was only an indifferent kindergarten. Europe and Asia were our principal schools.”