The Ravenous Brain: How the New Science of Consciousness Explains Our Insatiable Search for Meaning (29 page)
Authors: Daniel Bor

BOOK: The Ravenous Brain: How the New Science of Consciousness Explains Our Insatiable Search for Meaning
2.45Mb size Format: txt, pdf, ePub
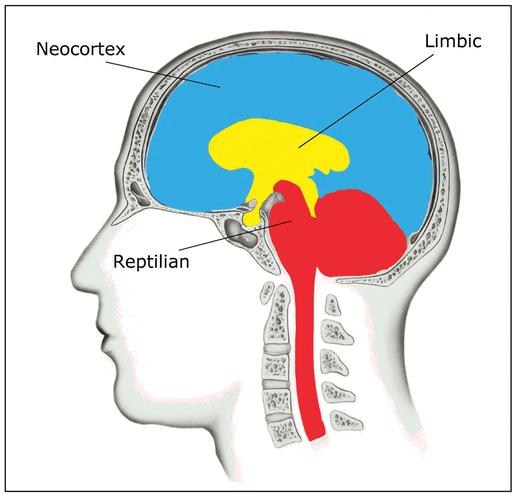
Figure 4
. The four lobes of the human brain.
. The four lobes of the human brain.
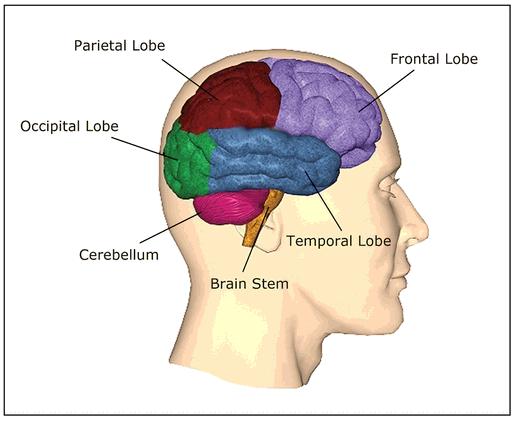
Figure 5
.
An example of change blindness. The two figures are repeatedly swapped, in between a blank grey screen, until the volunteer spots the blatant but unexpected change.
.
An example of change blindness. The two figures are repeatedly swapped, in between a blank grey screen, until the volunteer spots the blatant but unexpected change.
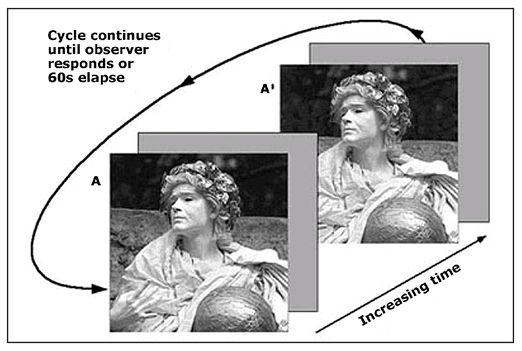
Figure 6
.
Examples of stimuli that induce repeated switches in visual perception. In the top example, involving binocular rivalry, a single mixed picture presents a face to one eye and a house to the other, because of red/green filtered glasses. The experience flips between only a face and only a house randomly over time. In the bottom two examples, we experience either only a candlestick or only two profile faces (left), and either only an old woman or only a young woman (right), and our experience again flips back and forth randomly over time.
.
Examples of stimuli that induce repeated switches in visual perception. In the top example, involving binocular rivalry, a single mixed picture presents a face to one eye and a house to the other, because of red/green filtered glasses. The experience flips between only a face and only a house randomly over time. In the bottom two examples, we experience either only a candlestick or only two profile faces (left), and either only an old woman or only a young woman (right), and our experience again flips back and forth randomly over time.
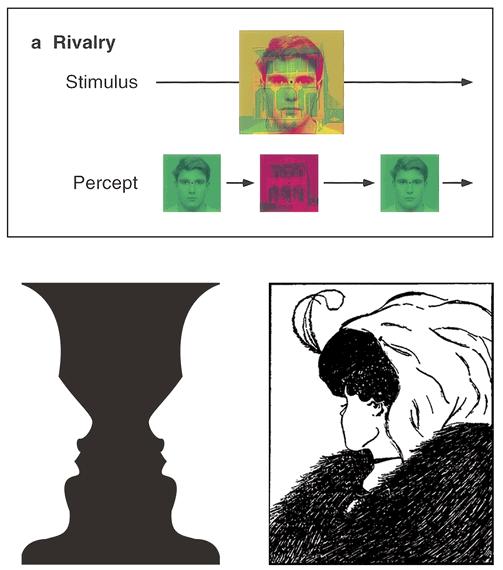
Figure 7
.
(a) Illustration of the three types of cup assembly. Strategy 1 merely involves putting one cup inside another, and not completing the puzzle. Strategy 2 involves putting the cups inside each other one at a time, and never moving a cup with another inside it. Although the puzzle can be completed in this way, it’s different from the demonstration that the experimenter showed the participant. Finally, there is strategy 3, which involves some level of hierarchy, as both the smaller and middle cups are moved simultaneously to fit into the larger cup. (b) Graphs to show dominant strategy used for infants and animals—strategy 1 in blue, 2 in magenta, and 3 in yellow.
.
(a) Illustration of the three types of cup assembly. Strategy 1 merely involves putting one cup inside another, and not completing the puzzle. Strategy 2 involves putting the cups inside each other one at a time, and never moving a cup with another inside it. Although the puzzle can be completed in this way, it’s different from the demonstration that the experimenter showed the participant. Finally, there is strategy 3, which involves some level of hierarchy, as both the smaller and middle cups are moved simultaneously to fit into the larger cup. (b) Graphs to show dominant strategy used for infants and animals—strategy 1 in blue, 2 in magenta, and 3 in yellow.
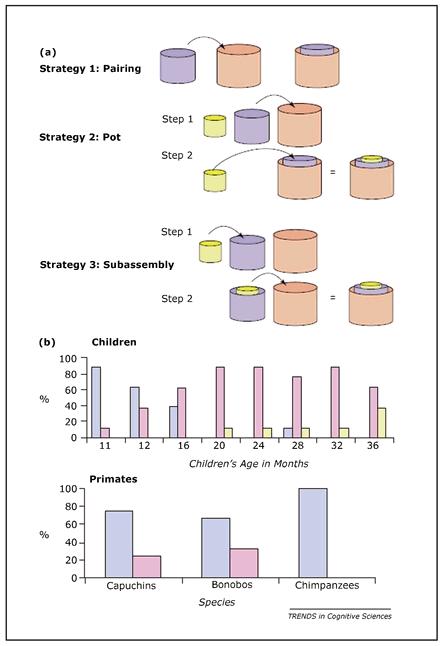
Figure 8
. In the fMRI scanner, normal volunteers see a 4 by 4 grid of red squares. Four of them blink blue in a sequence, which the volunteers have to remember for a few seconds. The left side is an example of a structured sequence, and the right side is an unstructured example.
. In the fMRI scanner, normal volunteers see a 4 by 4 grid of red squares. Four of them blink blue in a sequence, which the volunteers have to remember for a few seconds. The left side is an example of a structured sequence, and the right side is an unstructured example.
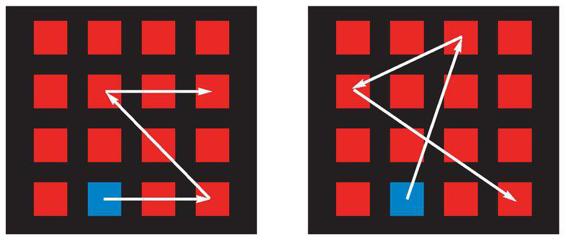
Figure 9
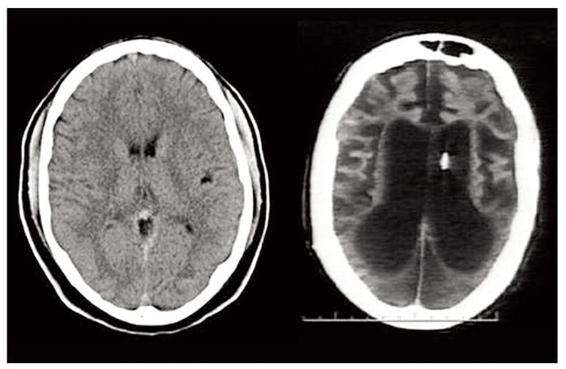
. A CT scan comparison of a normal brain, on the left, and Terri Schiavo’s brain, on the right.
. A CT scan comparison of a normal brain, on the left, and Terri Schiavo’s brain, on the right.
The study of how the brain creates consciousness involves two surprisingly disconnected wings. The first side has explicitly sought to discover the brain areas and neural mechanisms responsible for consciousness. But in parallel to this strand of research, a largely separate group of scientists has been revealing equally insightful discoveries about the neural recipes for our experiences, despite the fact that they rarely even mention consciousness. Instead, their topics of investigation include the neuroscience of working memory, attention, and chunking.
In this chapter, I will recount the direct, official evidence for how the brain generates our sense of awareness. But then I will show how the study of the neural underpinnings of attention, working memory, chunking, and related topics map closely onto the “official” set of consciousness data, providing further evidence for the importance of these processes to consciousness. In the final section of the chapter I will describe current neural theories of consciousness to provide deeper insights into its nature, especially when strengthened by a synthesis of the two experimental stories.
OPENING THE FLOODGATESProbably the simplest question to ask of the neuroscience of consciousness, and therefore a natural first point of attack, is whether the whole brain contributes to consciousness, or merely a few of its key areas. There is an unfortunate abundance of data on this question in the form of patients with damage to different brain regions. En masse, this population of patients has localized damage to every brain region there is. This allows us to know, for instance, that the cerebellum, part of the ancient reptilian section of the human brain, has little to do with awareness. Patients missing a cerebellum show no clear impairments of consciousness. For instance, one woman was born with almost no cerebellum in either hemisphere, but she was able to lead a relatively normal life, holding down a job at an electronics factory.
So certainly not all brain areas equally contribute to consciousness. The above example also partially answers another question from the list—about consciousness simply relating to number of neurons. If neuron count equated to levels of awareness, then the cerebellum, with perhaps 80 percent of the entire brain’s neurons, would be the most conscious part. That clearly isn’t the case.
In fact, the most critical region for consciousness is also one of the smallest and—in some ways—the least interesting, for the purposes of this book. This brain area is known as the reticular formation. It is part of the brain stem, the most primitive of brain regions and another component of the reptilian brain. The reticular formation controls the sleep-wake cycle through a complex set of subregions that each play a part in a chemical and neuronal cascade of activity. These actions allow us both to wake up and to enter different modes of sleep. For instance, when we dream, it is the reticular formation that pushes signals down to the spinal cord, actively paralyzing the rest of the body so that we don’t start actually crashing into walls when we’re dreaming we’re running around. Damage to the reticular formation is usually a pretty absolute business. Either the patient will die or he will be totally robbed of consciousness in a deep coma, unable to ever wake up.
But while we definitely need our reticular formation in order to be aware, it doesn’t follow that our consciousness actually occurs in this primitive brain area. My PC will simply not turn on without the power supply unit, but this box is the dumbest part of my computer, having nothing to do with the processing that gives me a functioning operating system.
A somewhat more relevant brain region is one of the main output regions for the reticular formation, the thalamus. It sits just above the brain stem, right in the center of the brain. This particular region is very special because it acts as a hub, the Grand Central Station of the brain. Its neurons receive inputs from, and send outputs to, almost every other brain region. Historically, it was viewed mainly as a sensory conduit, because the thalamus is usually the first port of call for the senses before the information is shunted to the cortex. For instance, for vision, our eyes report what they see via a thick information highway known as the optic tract, which flows into a part of the thalamus known as the lateral geniculate nucleus. This then shunts the information on to the primary visual cortex. More recently, though, the role of the thalamus has been considerably upgraded from passive relay. It is now known to be a sophisticated information-filtering and -organizing device and is thought to play a central role in consciousness.
Patients with extensive damage to the thalamus tend to enter a so-called vegetative state. This is distinguished from coma in that vegetative patients show signs of wakefulness—they may open their eyes more in the daytime, for instance. It’s as if their reticular formation is still partially carrying out its duty of waking up the brain, but there isn’t enough neural coherence for them actually to be aware of anything when they are awake. As the thalamus is a key player in generating consciousness, it’s natural that its functions have been discussed both in terms of information processing and attention: Along with helping with the flow of information between most regions of the brain, the thalamus helps to point our attention in various directions. This explains how those in a vegetative state with damage to their thalamus can appear to be awake, but lack any directed consciousness. Imagine a life where you were alert in some sense, but your attention was never directed toward any object, any goal, any thought—would we even call such a state conscious?
Other books
Charming (Exiled Book 3) by Victoria Danann
Vintage Munro by Alice Munro
Irresistible Force by D. D. Ayres
Not Quite Mine (Not Quite series) by Bybee, Catherine
Horse Named Dragon by Gertrude Chandler Warner
One Minute to Midnight by Amy Silver
Maxar (The Mating Games Book 4) by Catty Diva
WOLF: An Evil Dead MC Story (The Evil Dead MC Series Book 4) by Nicole James
One Hot Mess by Lois Greiman
One of Our Thursdays Is Missing by Jasper Fforde