The Seven Daughters of Eve (5 page)
Read The Seven Daughters of Eve Online
Authors: Bryan Sykes

The significance of all this to the thinking about European prehistory is that Mourant realized that having two Rhesus blood groups in a single population did not make any evolutionary sense. Even the simplest calculations showed that losing so many babies was not a stable arrangement. There was no problem if everybody had the same Rhesus type. It didn't matter whether this was Rhesus positive or Rhesus negative, just so long as it was all one or the other. It was only when there were people with different Rhesus types breeding together that these very serious problems arose. In the past, before blood transfusions and before the antibody treatment for Rhesus negative mothers, there must have been a lot of babies dying from haemolytic disease. This is a very heavy evolutionary burden, and the expected result of this unbalanced situation would be that one or other of the Rhesus blood groups would eventually disappear. And this is exactly what has happened â everywhere except in Europe. While the rest of the world is predominantly Rhesus positive, Europe stands out as having a very nearly equal frequency of both types. To Mourant, this was a signal that the population of Europe was a mixture that had not yet had time to settle down and eliminate one or other of the Rhesus types. His explanation was that modern Europe might be a relatively recent hybrid population of Rhesus positive arrivals from the Near East, probably the people who brought farming into Europe beginning about eight thousand years ago, and the descendants of an earlier Rhesus negative hunter-gathering people. But who were the Rhesus negatives?
Mourant came across the work of the French anthropologist H. V. Vallois, who described features of the skeletons of contemporary Basques as having more in common with fossil humans from about twenty thousand years ago than with modern people from other parts of Europe. Though this kind of comparison has since fallen into disrepute, it certainly catalysed Mourant's thinking. It was already known that Basques had by far the lowest frequency of blood group B of all the population groups in Europe. Could they be the ancient reservoir of Rhesus negative as well? In 1947 Mourant arranged to meet with two Basques who were in London attempting to form a provisional government and were keen to support any attempts to prove their genetic uniqueness. Like most Basques, they were supporters of the French Resistance and totally opposed to the fascist Franco regime in Spain. Both men provided blood samples and both were Rhesus negative. Through these contacts, Mourant typed a panel of French and Spanish Basques who turned out, as he had hoped, to have a very high frequency of Rhesus negatives, in fact the highest in the world. Mourant concluded from this that the Basques were descended from the original inhabitants of Europe, whereas all other Europeans were a mixture of originals and more recent arrivals, which he thought were the first farmers from the Near East.
From that moment, the Basques assumed the status of the population against which all ideas about European genetic prehistory were to be â and to a large extent still are â judged. The fact that they alone of all the west Europeans spoke a language which was unique in Europe, and did not belong to the Indo-European family which embraces all other languages of western Europe, only enhanced their special position.
The next leap forward came from the mathematical amalgamation of the vast amount of data that had accumulated from decades of research on individual systems like the different blood groups. This was accomplished by the man who has dominated the field for the past thirty years, Luigi Luca Cavalli-Sforza. We will meet him again later. Cavalli-Sforza, working with the Cambridge statistician Anthony Edwards, achieved this amalgamation using the earliest punched-card computing machines. By averaging across several genetic systems at once they managed to eliminate most of the bizarre and counter-intuitive conclusions that had discredited the anthropological applications of blood groups when they were worked on one at a time. The weakness of using just a single system was that two populations, like the Russians and the Malagasy, could end up with the same gene frequency just by chance rather than because of a common ancestry. This was far less likely to happen if several genes were compared, because the impact of a misleading result from one of them would be diluted out by the effect of the others. There were to be no more Russian invasions of Madagascar. None the less, the underlying principle remained the same. In an evolutionary sense, populations with similar gene frequencies were more likely to be closely related to each other than populations whose gene frequencies were very different.
Anthony Edwards explained his thinking in an ingenious article in
New Scientist
in 1965. He imagines a tribe that carries with it a pole along which are arrayed 100 discs which are either black or white. Every year, one disc, chosen at random, is changed to the other colour. When the tribe splits into two groups, each group takes with it a copy of the pole with the discs in their current order. The following year they each make one of the random changes to the discs. The next year they make another, the next year another and so on, continuing the custom of one random change every year. Since the changes they make are completely random, the order of the discs on the two poles becomes more and more dissimilar as each year passes. It follows that if you were to look at the poles carried by the two tribes you could estimate how long ago, in a relative sense, they separated from each other by the differences in the order of the black and white discs. Providing an absolute date was very difficult from the gene frequency data alone, but the comparative separation between the two tribes, known as the
genetic distance
, was a useful measure of their common ancestry. The bigger the genetic distance between them, the longer they had spent apart.
This was a clever image of the process of genetic change, called
genetic drift
, brought about by the random survival and extinction of genes as they pass from one generation to the next. This process leads to bigger and bigger differences in the frequencies of genes as time passes. Just like the order of discs in Edwards' analogy, gene frequencies can be used to backtrack and work out how long ago two groups of people were once together as a single population. These groups could be villages, tribes or whole populations, and there is no limit to the number of groups that can be analysed in this way. If you do it for the whole world, the outcome is a diagram like Figure 1 overleaf.
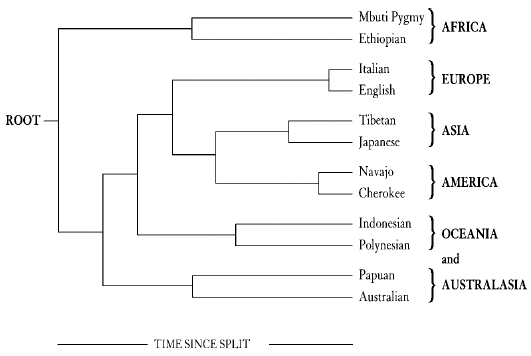
Figure 1
Along the right-hand side we have several âpopulations' (I have picked two examples from each continent) and along the bottom is the genetic distance/time axis. This is what is called a population tree where the lines trace, from left to right, the estimated order in which âpopulations' evolved and split from one another, as reconstructed from the assimilated frequencies of many different genes. At first glance, many of the groupings look quite sensible. The two European populations, the English and the Italians, are close together on two short âbranches' of the tree. The two native American tribes are connected together with their closest relatives in Asia, as we would expect if the first Americans crossed the Bering land bridge from Siberia to Alaska. The two populations from Africa are on a different branch from the rest of the world, which correctly emphasizes that continent's great antiquity as the cradle of human evolution. This is a much more sensible-looking tree than can be drawn from the First World War blood group data which, as well as allying Russia and Madagascar, entirely missed the importance of Africa. The reason for this, as noted earlier, is that the odd quirks that arose by chance with a single system, like the ABO blood groups, get ironed out by amalgamating the results from several different genes.
Edwards acknowledged that âThe resultant evolutionary trees will certainly not provide the last word on human evolution,' and offered the diagrams as a way of providing the genetic information in an understandable form. Unfortunately, the population trees first drawn with this admirable and modest intention were over-interpreted and became a source of contention. Among the several reasons for this is just the way they appear. They do look as if they are real evolutionary trees and have often been portrayed as exactly that. They could only be true evolutionary trees if human evolution really were a succession of population fissions along the lines of the splits that Edwards explains in his metaphor of the tribes with their poles and discs. Then and only then would the nodes, the points on the tree from which two lines diverge, represent a real entity. These would be the populations that existed before the splits, the proto-populations. But is that what really happened in human evolution? For instance, in the European part of the tree, was there ever such a thing as the proto-Anglo-Italian population which divided, never to meet again, and became the modern inhabitants of England and Italy? That might have been the case if the English and Italians became two different species as soon as they split and could never interbreed again. But they can, and they do, and they always have done. As we will discover later in the book, humans just did not evolve like this.
Perhaps the most serious objection to these trees is that their construction demands that the things at the end of the trees, the populations, be objectively defined. This process in itself segregates people into groups in ways that can tend to perpetuate racial classifications. It gives some sort of overall genetic number to something that does not really exist. There are certainly
people
who live in Japan and Tibet, but there is no genetic meaning to
the population
of Tibet or Japan, taken as a whole. As this book will show, objectively defined races simply do not exist. Even Arthur Mourant realized that fact nearly fifty years ago, when he wrote: âRather does a study of blood groups show a heterogeneity in the proudest nation and support the view that the races of the present day are but temporary integrations in the constant process ofâ¦mixing that marks the history of every living species.' The temptation to classify the human species into categories which have no objective basis is an inevitable but regrettable consequence of the gene frequency system when it is taken too far. For several years the study of human genetics got firmly bogged down in the intellectually pointless (and morally dangerous) morass of constructing ever more detailed classifications of human population groups.
Fortunately, there was a way out of this impasse. The break-out came with the publication of a scientific paper in
Nature
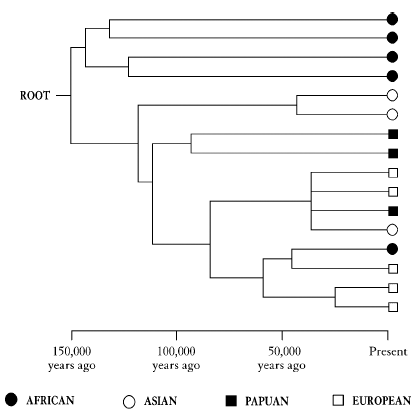
in January 1987 by the veteran US evolutionary biochemist, the late Allan Wilson, and two of his students, Rebecca Cann and Mark Stoneking, entitled âMitochondrial DNA and human evolution'. The centrepiece of this article was a diagram which bears a superficial resemblance to the trees I have just been criticizing. I have reproduced a small section of it here in Figure 2, with only sixteen individuals instead of the 134 in the original paper.
It is indeed an evolutionary tree; but this time the diagram means something. On the right of the tree the symbols at the tips of the branches represent not populations but the sixteen
individuals
that I have selected to illustrate the point, sixteen people from four different parts of the world: Africans, Asians, Europeans and Papuans from New Guinea. The first improvement over the other trees is that, unlike
populations
, there is no argument about whether
people
exist or not. They clearly do. The other improvement is that the nodes on the tree are also real people and not some hypothetical concept like a âproto-population'. They represent the last common ancestors of the two people who branch off from that point. The lines that connect the sixteen people on the diagram are drawn to reflect genetic differences between them in one very special gene called mitochondrial DNA whose unusual and useful properties I will introduce shortly. For reasons I shall explain in the next chapter, if two people have very similar mitochondrial DNA then they are more closely related, with respect to this gene, than two people with very different mitochondrial DNA. They have a common ancestor who lived more recently in the past, and so are joined by shorter branches on the diagram. People with very different mitochondrial DNA share a more remote common ancestor and are linked by longer branches.