Bully for Brontosaurus (12 page)
Read Bully for Brontosaurus Online
Authors: Stephen Jay Gould

The alternative interpretation holds that kiwis are phyletic dwarfs, evolved from a lineage of much larger birds. Since these large ancestors laid big eggs appropriate to their body size, kiwis just never (or only slightly) reduced the size of their eggs as their bodies decreased greatly in bulk. In other words, kiwi eggs never became unusually large; kiwi bodies got small—and these statements are not equivalent, just as we know that an obese man is not short for his weight, despite the old jest.
(Such a hypothesis is not anti-adaptationist in the sense that maintenance of a large egg as size decreases—and in the face of energetic and biochemical costs imposed by such a whopping contribution to the next generation—may well require a direct boost from natural selection to prevent an otherwise advantageous decrease more in keeping with life at Colonel Sanders’s favorite size. Still, there is a world of difference between retaining something you already have, and first developed for other reasons [in this case simple appropriateness for large body size], and actively evolving such a unique and cumbersome structure for some special benefit.)
Calder’s interpretation might seem forced or farfetched but for the outstanding fact of taxonomy and biogeography cited as the introduction to this essay. Moas are the closest cousins of kiwis, and most moas were very large birds. “Is the kiwi perhaps a shrunken moa?” Calder asks. Unfortunately, all moa fossils lie in rocks of a geological yesterday, and kiwi fossils are entirely unknown—so we have no direct evidence about the size of ancestral kiwis. Still, I believe that all the inferential data support Calder’s alternative hypothesis for the great size of kiwi eggs—a “structural” or “historical” explanation if you will, not a conventional account based on natural selection for immediate advantages.
Although the best argument for viewing kiwis as much smaller than their ancestors must be the large size of their closest moa cousins, Calder has also developed a quirky and intriguing speculation to support the dwarfed status of kiwis. (I hasten to point out that neither of these arguments amounts to more than a reasonable conjecture. All evidence can be interpreted in other ways. Both moas and kiwis, for example, might have evolved from a kiwi-sized common ancestor, with moas enlarging later. Still, since the kiwi is the smallest of all ratites—a runt among ostriches, rheas, emus, and cassowaries—its decrease seems more probable than moa increase. But we will not know until we have direct evidence of fossil ancestry.)
Calder notes that in many respects, some rather curious, kiwis have adopted forms and lifestyles generally associated with mammals, not birds. Kiwis, for example, are unique among birds in retaining ovaries on both sides (the right ovary degenerates in all other birds)—and eggs alternate between sides, as in mammals. The seventy-to eighty-four-day incubation period matches the eighty-day pregnancy expected for a mammal of kiwi body size, not the forty-four days predicted for birds of this weight. Calder continues: “When one adds to this list, the kiwi’s burrow habit, its furlike body feathers, and its nocturnal foraging highly dependent on its sense of smell, the evidence for convergence seems overpowering.” Of course, this conjunction of traits could be fortuitous and each might mean something quite unmammalian to a kiwi, but the argument does gain strength when we remember that no terrestrial mammals reached New Zealand, and that the success of many introduced species indicates a hospitable environment for any creature that could exploit a mammalian way of life.
You will be wondering what these similarities with mammals could possibly mean for my key claim that kiwis are probably descendants of much larger birds. After all, mammals are superior, noble, and large. But they aren’t. The original and quintessential mammalian way of life (still exploited by a majority of species) is secretive, furtive, nocturnal, smell-oriented in a non-visual world—and, above all, small. Remember that for two-thirds of their geological history, all mammals were little creatures living in the interstices of a world ruled by dinosaurs. If a large bird converged upon a basically mammalian lifestyle in the absence of “proper” inhabitants as a result of geographic isolation,
decrease
in size would probably be a first and best step.
Perhaps I have convinced you that kiwis probably decreased in size during their evolution. But why should this dwarfing help to explain their large eggs? Why didn’t egg size just keep pace with body size as kiwis scaled down? We now come to the strong evidence of the case.
The study of changes in form and proportion as organisms increase or decrease in size is called allometry. It has been a popular and fruitful subject in evolutionary research since Julian Huxley’s pioneering work of the 1920s. One of Huxley’s own classic studies (
Journal of the Linnaean Society of London
, 1927) bore the title: “On the Relation between Egg-weight and Body-weight in Birds.” Huxley found that if you plot one point for each species on the hummingbird-to-moa curve for egg weight versus body weight, relative egg size decreases in an even and predictable way. The eggs of large birds, he found, are absolutely larger, but relatively smaller in proportion to body weight, than those of small birds.
Huxley’s work has since been extended several times with more voluminous and consistent data. In the two best studies that I know, Samuel Brody (in his masterful compendium,
Bioenergetics and Growth
, 1945) calculated a slope of 0.73, while H. Rahn, C. V. Paganelli, and A. Ar (1975), with even more data from some 800 species, derived a similar value of 0.67. This means that as birds increase in body weight, egg weight enlarges only about two-thirds as fast. Conversely, as birds decrease in size, egg weight diminishes more slowly—so little birds have relatively heavy eggs.
This promising datum will not, however, explain the kiwi’s outsized egg, for the two-thirds slope represents the general standard for all birds. Kiwi eggs are huge compared with the
expected
egg weight for a bird of kiwi body weight along this standard curve.
But the literature of allometry has also yielded a generality that will, I think, explain the kiwi’s massive egg. The two-thirds slope of the egg weight/body weight curve represents a type of allometry technically called interspecific scaling—that is, you plot one point for each species in a related group of organisms and attempt to establish the characteristic change of proportion along a gradient of increasing size. (These curves are popularly called mouse-to-elephant for relationships among mammals—hence my designation hummingbird-to-moa for birds.) Allometricians have established hundreds of interspecific curves for birds and mammals.
Another kind of allometry is called intraspecific scaling. Here you plot one point for each individual among adults of varying body weights within a single species—the Tom Thumb-to-Manute Bol curve for human males, if you will. Since the similarity of these technical terms—interspecific and intraspecific—is so confusing, I shall call them, instead, among-species (for mouse-to-elephant) and within-species (for Thumb-to-Bol).
As an important generality in allometric studies, within-species curves usually have a substantially lower slope than among-species curves for the same property. For example (and in our best-studied case), the mouse-to-elephant curve for brain weight versus body weight in mammals has a slope of about two-thirds (as does the egg weight/body weight curve for birds). But the within-species curve from small to large adults of a single species, while varying from one group to another, almost always has a much lower slope in the range of 0.2 to 0.4. In other words, while brains increase about two-thirds as fast as bodies among species (implying that large mammals have relatively small brains), brains only increase about one-fifth to two-fifths as fast as bodies when we move from small to large adults within a single mammalian species.
Such a regularity, if it applied to egg weight as well, could resolve the kiwi paradox—if kiwis evolved from larger ancestors. Suppose that kiwi forebears start at moa size. By the hummingbird-to-moa among-species standard, egg size should decrease along the two-thirds slope. But suppose that natural selection is operating to favor small adults within a population. If the within-species curve for egg weight had a slope much lower than two-thirds, then size decrease by continued selection of small adults might produce a new species with outsized eggs well above the two-thirds slope, and therefore well above the expected weight for a bird of this reduced size. (Quantitative arguments like this are always easier to grasp by picture than by words—and a glance at the accompanying graph should resolve any confusion.)
But what is the expected within-species relationship for egg weight? Is the shape of the curve low, as for brain weight, thus affirming my conjecture? I reached for my well-worn copy of Brody’s unparalleled compendium and found that for adults of domestic fowl, egg weight increases not two-thirds as fast, but only 15 percent as fast as body weight! (Brody uses this fact to argue that small hens are usually better than large, so long as egg production remains the same—for egg size diminishes very little with a large decrease in body mass, and the small loss in egg volume is more than compensated by large decreases in feeding costs.)
The same argument might apply to kiwis. As a poultryman might choose small hens for minimal decrease in egg size with maximal decline in body weight, natural selection for smaller adults might markedly decrease the average body weight within a species with very little accompanying reduction in egg weight.
I believe that this general argument, applied to kiwis, may be defended on three strong grounds. First, as stated above, a general finding in allometric studies teaches us that within-species slopes for adults of one species are usually much lower than among-species slopes along mouse-to-elephant curves. Thus, any evolution of decreasing size along the within-species curve should produce a dwarfed descendant with more of the particular item being measured than an average nondwarfed species at the same body weight. Second, we have actual data, for domestic poultry at least, indicating that the within-species curve does have a substantially lower slope than the hummingbird-to-moa curve for our crucial measure of egg weight.
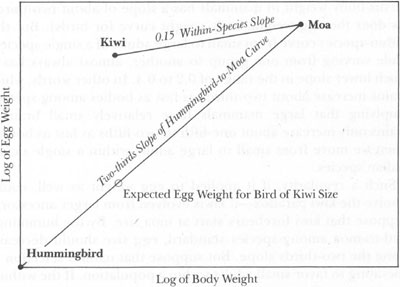
Proposed allometric explanation for the large egg of the kiwi. The kiwi probably evolved from a much larger bird by backing down the very shallow within-species slope (upper line). Most birds arrange themselves on the standard hummingbird-to-moa curve with its steeper slope (lower line). Therefore, a kiwi has a much heavier egg than predicted for a bird of its body size.
BEN GAMIT. ADAPTED FROM JOE LEMONNIER. COURTESY OF
NATURAL HISTORY
.
Third, I have studied many cases of dwarfism, and I believe we can state as a general phenomenon—rooted in the first point above—that decline in body size often far outstrips decrease in many particular features. Dwarfs, in several respects, always seem to have much more of certain body parts than related nondwarfed species of the same body size. For example, I once studied tooth size in three species of dwarfed hippos (two fossil and the modern Liberian pygmy)—and found their molar teeth substantially larger, for each of three separate evolutionary events, than expected values for related hoofed mammals at their body size (
American Zoologist
, 1975).
In another example, the talapoin, a dwarfed relative of the rhesus monkey, has the largest relative brain weight among monkeys. Since within-species brain curves have substantially lower slopes than the two-thirds value for the marmoset-to-baboon curve, evolution to smaller size by backing down the within-species curve would yield a dwarf with a far larger brain than an ordinary monkey at the same body size.
Put all this together and a resolution fairly jumps at you for kiwis. Their enormous eggs require no special explanation if kiwis have evolved by marked decrease in size. Kiwi eggs exhibit the weight expected for backing down the within-species curve if natural selection operates only to decrease body size and no other factor intervenes to favor an active reduction in egg size—as we might anticipate in New Zealand, this easy land of no natural predators, where a female might waddle without fear as an enormous egg distends her abdomen during passage down the oviduct.
In this interpretation, if you ask me why kiwi eggs are so large, I reply, “Because kiwis are dwarfed descendants of larger birds, and just followed ordinary principles of scaling in their evolution.” This answer differs sharply from the conventional form of evolutionary explanation: “Because these big eggs are good for something now, and natural selection favored them.”
My answer will also strike many people as deeply unsatisfactory. It provides a reason rooted in history, pure and simple (with a bit of scaling theory thrown in)—kiwis are as they are because their ancestors were as they were. Don’t we want answers that invoke general laws of nature rather than particular contingencies of history?
I would reply that my resolution is quite satisfactory, that evolutionary arguments are often properly resolved by such historical statements, and that we would do well to understand this important and neglected principle of reasoning—for we might save ourselves many a stumble in trying to apply preferred, but inappropriate, styles of explanation to situations encountered again and again in our daily lives.