Bully for Brontosaurus (16 page)
Read Bully for Brontosaurus Online
Authors: Stephen Jay Gould

In preparing this essay, I spent several days reading the classical literature on the evolution of insect flight—and emerged with a deeper understanding of just how difficult Darwin’s principle of functional shift can be, even for professionals. Most of the literature hasn’t even made the first step of applying functional shift at all, not to mention the later reform of substituting direct evidence for verbal speculation. Most reconstructions are still trying to explain the incipient stages of insect wings as somehow involved in airborne performance from the start—not for flapping flight, of course, but still for some aspect of motion aloft rather than, as Darwin’s principle would suggest, for some quite different function.
To appreciate the dilemma of such a position (so well grasped by Mivart more than 100 years ago), consider just one recent study (probably the best and most widely cited) and the logical quandaries that a claim of functional continuity entails. In 1964, J. W. Flower presented aerodynamic arguments for wings evolved from tiniest rudiment to elaborate final form in the interest of airborne motion. Flower argues, supporting an orthodox view, that wings evolved from tiny outgrowths of the body used for gliding prior to elaboration for sustained flight. But Flower recognizes that these incipient structures must themselves evolve from antecedents too small to function as gliding planes. What could these very first, slight outgrowths of the body be for? Ignoring Darwin’s principle of functional shift, Flower searches for an aerodynamic meaning even at this very outset. He tries to test two suggestions: E. H. Hinton’s argument that initial outgrowths served for “attitude control,” permitting a falling insect to land in a suitable position for quick escape from predators; and a proposal of the great British entomologist Sir Vincent Wigglesworth (wonderful name for an insect man, I always thought) that such first stages might act as stabilizing or controlling devices during takeoff in small, passively aerial insects.
Flower proceeded by performing aerodynamic calculations on consequences of incipient wings for simple body shapes when dropped—and he quickly argued himself into an inextricable logical corner. He found, first of all, that tiny outgrowths might help, as Wigglesworth, Hinton, and others had suggested. But the argument foundered on another observation: The same advantages could be gained far more easily and effectively by another, readily available alternative route—evolution to small size (where increased surface/volume ratios retard falling and enhance the probability of takeoff). Flower then realized that he would have to specify a reasonably large body size for incipient wings to have any aerodynamic effect. But he then encountered another problem: At such sizes, legs work just as well as, if not better than, proto-wings for any suggested aerodynamic function. Flower admitted:
The first conclusion to be drawn from these calculations is that the selective pressure in small insects is towards smaller insects, which would have no reason to evolve wings.
I would have stopped and searched elsewhere (in Darwin’s principle of functional shift) at this point, but Flower bravely continued along an improbable path:
The main conclusions, however, are that attitude control of insects would be by the use of legs or by very small changes in body shape [
i.e.
, by evolving small outgrowths, or proto-wings].
Flower, in short, never considered an alternative to his assumption of functional continuity based upon some aspect of aerial locomotion. He concluded:
At first they [proto-wings] would affect attitude; later they could increase to a larger size and act as a true wing, providing lift in their own right. Eventually they could move, giving the insect greater maneuverability during descent, and finally they could “flap,” achieving sustained flight.
As an alternative to such speculative reconstructions that work, in their own terms, only by uncomfortable special pleading, may I suggest Darwin’s old principle of functional shift (preadaptation—ugh—for something else).
The physiological literature contains voluminous testimony to the thermodynamic efficiency of modern insect wings: in presenting, for example, a large surface area to the sun for quick heating (see B. Heinrich, 1981). If wings can perform this subsidiary function now, why not suspect thermoregulation as a primary role at the outset? M. M. Douglas (1981), for example, showed that, in
Colias
butterflies, only the basal one-third of the wing operates in thermoregulation—an area approximately equal to the thoracic lobes (proto-wings) of fossil insects considered ancestral to modern forms.
Douglas then cut down some
Colias
wings to the actual size of these fossil ancestral lobes and found that insects so bedecked showed a 55 percent greater increase in body temperature than bodies deprived of wings entirely. These manufactured proto-wings measured 5 by 3 millimeters on a body 15 millimeters long. Finally, Douglas determined that no further thermoregulatory advantage could be gained by wings longer than 10 millimeters on a 15-millimeter body.
Kingsolver and Koehl performed a host of elaborate and elegant experiments to support a thermoregulatory origin of insect proto-wings. As with so many examples of excellent science producing clear and interesting outcomes, the results can be summarized briefly and cleanly.
Kingsolver and Koehl begin by tabulating all the aerodynamic hypotheses usually presented in the literature as purely verbal speculations. They arrange these proposals of functional continuity (the explanations that do not follow Darwin’s solution of Mivart’s dilemma) into three basic categories: proto-wings for gliding (aerofoils for steady-state motion), for parachuting (slowing the rate of descent in a falling insect), and attitude stability (helping an insect to land right side up). They then transcended the purely verbal tradition by developing aerodynamic equations for exactly how proto-wings should help an insect under these three hypotheses of continuity in adaptation (increasing the lift/drag ratio as the major boost to gliding, increasing drag to slow the descent rate in parachuting, measuring the moment about the body axis produced by wings for the hypothesis of attitude stability).
They then constructed insect models made of wire, epoxy, and other appropriate materials to match the sizes and body shapes of flying and nonflying forms among early insect fossils. To these models, they attached wings (made of copper wire enclosing thin, plastic membranes) of various lengths and measured the actual aerodynamic effects for properties predicted by various hypotheses of functional continuity. The results of many experiments in wind tunnels are consistent and consonant: Aerodynamic benefits begin for wings above a certain size, and they increase as wings get larger. But at the small sizes of insect proto-wings, aerodynamic advantages are absent or insignificant and do not increase with growing wing length. These results are independent of body shape, wind velocity, presence or placement of legs, and mounting position of wings. In other words, large wings work well and larger wings work better—but small wings (at the undoubted sizes of Mivart’s troubling incipient stages) provide no aerodynamic edge.
Kingsolver and Koehl then tested their models for thermoregulatory effects, constructing wings from two materials with different thermal conductivities (construction paper and aluminum foil) and measuring the increased temperature of bodies supplied with wings of various lengths versus wingless models. They achieved results symmetrically opposite to the aerodynamic experiments. For thermoregulation, wings work well at the smallest sizes, with benefits increasing as the wing grows. However, beyond a measured length, further increase of the wing confers no additional effect. Kingsolver and Koehl conclude:
At any body size, there is a relative wing length above which there is no additional thermal effect, and below which there is no significant aerodynamic effect.
The accompanying chart illustrates these combined results. Note how the thermoregulatory effect of excess body temperature due to wings (solid line) increases rapidly at small wing sizes but not at all above an intermediate wing length. Conversely, the aerodynamic effect of lift/drag ratio does not increase at all until intermediate wing length, but grows rapidly thereafter.
We could not hope for a more elegant experimental confirmation of Darwin’s solution to Mivart’s challenge. Kingsolver and Koehl have actually measured the functional shift by showing that incipient wings aid thermoregulation but provide no aerodynamic benefit—while larger wings provide no further thermoregulatory oomph but initiate aerodynamic advantage and increase the benefits steadily thereafter. The crucial intermediate wing length, where thermoregulatory gain ceases and aerodynamic benefits begin, represents a domain of functional shift, as aerodynamic advantages pick up the relay from waning thermoregulation to continue the evolutionary race to increasing wing size.
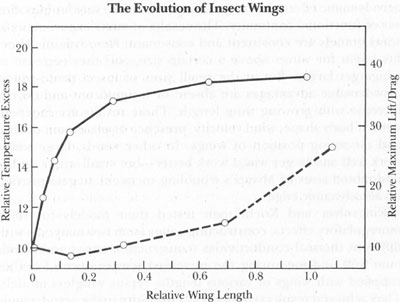
The thermoregulatory (upper curve) and aerodynamic (lower curve) advantages for increasing wing length in insects. Note that thermodynamic benefits accrue rapidly when the wing is very small (too small for flight), but scarcely increase at all for wings of larger size. Aerodynamic advantages, on the other hand, are insignificant for small size, but increase rapidly at larger wing dimensions, just as the thermodynamic benefits cease.
BEN GAMIT. ADAPTED FROM JOE LEMONNIER. COURTESY OF
NATURAL HISTORY
.
But what might push an insect across the transition? Why reach this crucial domain at all? If wings originally worked primarily for thermoregulation, why not just stop as the length of maximum benefit approached? Here, Kingsolver and Koehl present an interesting speculation based on another aspect of their data. They found that the domain of transition between thermal and aerial effects varied systematically with body size: The larger the body, the sooner the transition (in terms of relative wing length). For a body 2 centimeters long, the transition occurred with wings 40 to 60 percent of body length; but a 10-centimeter body switches to aerodynamic advantage at only 10 percent of body length.
Now suppose that incipient ancestral wings worked primarily for thermoregulation and had reached a stable, optimum size for greatest benefit. Natural selection would not favor larger wings and a transition to the available domain of aerodynamic advantage. But if body size increased for other reasons, an insect might reach the realm of aerial effects simply by growing larger, without any accompanying change of body shape or relative wing length.
We often think, naively, that size itself should make no profound difference. Why should just more of the same have any major effect beyond simple accumulation? Surely, any major improvement or alteration must require an extensive and explicit redesign, a complex reordering of parts with invention of new items.
Nature does not always match our faulty intuitions. Complex objects often display the interesting and paradoxical property of major effect for apparently trifling input. Internal complexity can translate a simple quantitative change into a wondrous alteration of quality. Perhaps that greatest and most effective of all evolutionary inventions, the origin of human consciousness, required little more than an increase of brain power to a level where internal connections became rich and varied enough to force this seminal transition. The story may be much more complex, but we have no proof that it must be.
Voltaire quipped that “God is always for the big battalions.” More is not always better, but more can be very different.
WHEN ASTA
the fox terrier exhumed the body of the Thin Man, his delightfully tipsy detective master, Nick Charles, exclaimed, “You’re not a terrier; you’re a police dog” (
The Thin Man
, MGM 1934 original with William Powell and Myrna Loy). May I now generalize for Asia’s breed in the case of the telltale textbook.
The wisdom of our culture abounds with mottoes that instruct us to acknowledge the faults within ourselves before we criticize the failings of others. These words range from clichés about pots and kettles to various sayings of Jesus: “And why beholdest thou the mote that is in thy brother’s eye, but perceivest not the beam that is in thine own eye?” (Luke 6:41); “He that is without sin among you, let him first cast a stone at her” (John 8:7). I shall follow this wisdom by exposing my own profession in trying to express what I find so desperately wrong about the basic tool of American teaching, the textbook.
In March 1987, I spent several hours in the exhibit hall of the National Science Teachers Association convention in Washington, D.C. There I made an informal, but reasonably complete, survey of evolution as treated (if at all) in major high-school science textbooks. I did find some evidence of adulteration, pussyfooting, and other forms of capitulation to creationist pressure. One book,
Life Science
, by L. K. Bierer, K. F. Liem, and E. P. Silberstein (Heath, 1987), in an accommodation that at least makes you laugh while you weep for lost integrity in education, qualifies every statement about the ages of fossils—usually in the most barbarous of English constructions, the passive infinitive. We discover that trilobites are “believed to have lived 500–600 million years ago,” while frozen mammoths are “thought to have roamed the tundra 22,000 years ago.” But of one poor bird, we learn with terrible finality, “There are no more dodoes living today.” Their extinction occurred within the bounds of biblical literalism and need not be hedged.
But I was surprised and pleased to note that most books contained material at reasonable length about evolution, and with no explicit signs of tampering to appease creationists. Sins imposed by others were minimal. But I then found the beam in our own eye and became, if anything, more distressed than by any capitulation to the yahoos. The problem does not lie in what others are doing to us, but in what we are doing to ourselves. In book after book, the evolution section is virtually cloned. Almost all authors treat the same topics, usually in the same sequence, and often with illustrations changed only enough to avoid suits for plagiarism. Obviously, authors of textbooks are copying material on a massive scale and passing along to students an ill-considered and virtually Xeroxed version with a rationale lost in the mists of time.
Just two months after making this depressing observation, I read Diane B. Paul’s fascinating article “The Nine Lives of Discredited Data” (
The Sciences
, May 1987). Paul analyzed the sections on heritability of IQ from twenty-eight textbooks on introductory genetics published between 1978 and 1984. She paid particular attention to their treatment of Sir Cyril Burt’s data on identical twins raised separately. We now know that these “studies” represent one of the most striking cases of fraud in twentieth-century science—for Burt invented both data and co-workers. His sad story had been well publicized, and all authors of texts published since 1978 surely knew that Burt’s data had been discredited and could not be used. Several texts even included discussions of the Burt scandal as a warning about caution and scrutiny in science.
But Paul then found that nearly half these books continued to cite and use Burt’s data, probably unconsciously. Of nineteen textbooks that devoted more than a paragraph to the subject of genetics and IQ, eleven based their conclusions about high heritability on a review article published in
Science
in 1963. This review featured a figure that ten of these textbooks reproduced either directly or in slightly altered and simplified form. This figure includes, as a prominent feature, the results of Sir Cyril Burt (not yet suspect in 1963). We must conclude that the authors of these texts either had not read the 1963 article carefully or had not consulted it at all. Paul infers (correctly, I am sure) that this carelessness arises because authors of textbooks copy from other texts and often do not read original sources. How else to explain the several books that discussed the Burt scandal explicitly and then, unbeknownst to their authors, used the same discredited data in a figure?
Paul argues that the increasing commercialization of textbooks has engendered this virtual cloning of contents. Textbook publishing is a big business, replete with market surveys, fancy art programs, and subsidiary materials in the form of slide sets, teachers’ guides, even test-making and grading services. The actual text of the book can become secondary and standardized; any departure from a conventional set of topics could derail an entire industry of supporting materials. Teachers are also locked into a largely set curriculum based on this flood of accoutrements. Paul concludes: “Today’s textbooks are thicker, slicker, more elaborate, and more expensive than they used to be. They are also more alike. Indeed, many are virtual clones, both stylistic and substantive, of a market leader.”
The marketplace rules. Most publishing houses are now owned by conglomerates—CBS, Raytheon, and Coca-Cola among them—with managers who never raise their eyes from the financial bottom line, know little or nothing about books, and view the publishing arm of their diversified empire as but one more item for the ultimate balance. I received a dramatic reminder of this trend last week when I looked at the back cover of my score for Mozart’s
Coronation Mass
, now under rehearsal in my chorus. It read: “Kalmus Score. Belwin Mills Publishing Company, distributed by Columbia Pictures Publication, a unit of the Coca-Cola Company.” I don’t say that Bill Cosby or Michael Jackson or whoever advertises the stuff doesn’t like Mozart; I merely suspect that Don Giovanni can’t be high on the executive agenda when the big boys must worry about such really important issues as whether or not to market Cherry Coke (a resounding “yes” vote from this old New York soda fountain junkie).
Paul quotes a leading industry analyst from the 1984
Book Publishing Annual
. Future textbooks, the analyst argues, will have “more elaborate designs and greater use of color…. The ancillary packages will become more comprehensive…. New, more aggressive marketing plans will be needed just to maintain a company’s position. The quality of marketing will make the difference.” Do note the conspicuous absence of any mention whatsoever about the quality of the text itself.
Paul is obviously correct in arguing that this tendency to cloning has accelerated remarkably as concerns of the market overwhelm scholarly criteria in the composition of textbooks. But I believe that the basic tendency has always been present and has a human as well as a corporate face. Independent thought has always been more difficult than borrowing, and authors of textbooks have almost always taken the easier way out. Of course I have no objection to the similar recording of information by textbooks. No author can know all the byways of a profession, and all must therefore rely on written sources for areas not enlightened by personal expertise. I speak instead of the thoughtless, senseless, and often false copying of phrase, anecdote, style of argument, and sequence of topics that perpetuates itself by degraded repetition from text to text and thereby loses its anchor in nature.
I present an example that may seem tiny and peripheral in import. Nevertheless, and perhaps paradoxically, such cases provide our best evidence for thoughtless copying. When a truly important and well-known fact graces several texts in the same form, we cannot know whether it has been copied from previous sources or independently extracted from any expert’s general knowledge. But when a quirky little senseless item attains the frequency of the proverbial bad penny, copying from text to text is the only reasonable interpretation. There is no other source. My method is no different from the standard technique of bibliographic scholars, who establish lineages of texts by tracing errors (particularly for documents spread by copyists before the invention of printing).
When textbooks choose to illustrate evolution with an example from the fossil record, they almost invariably trot out that greatest warhorse among case studies—the history of horses themselves (see the next essay in this section for fallacies of the usual tale). The standard story begins with an animal informally called
Eohippus
(the dawn horse), or more properly,
Hyracotherium
. Since evolutionary increase in size is a major component of the traditional tale, all texts report the diminutive stature of ancestral
Hyracotherium
. A few give actual estimates or measurements, but most rely upon a simile with some modern organism. For years, I have been much amused (and mildly bothered) that the great majority of texts report
Hyracotherium
as “like a fox-terrier” in size. I was jolted into action when I found myself writing the same line, and then stopped. “Wait a minute,” said my inner voice, “beyond some vague memories of Asta last time I watched a Thin Man movie, I haven’t the slightest idea what a fox terrier is. I can’t believe that the community of textbook authors includes only dog fanciers—so if I don’t know, I’ll bet most of them don’t either.” Clearly, the classic line has been copied from text to text. Where did it begin? What has been its history? Is the statement even correct?
My immediate spur to action came from a most welcome and unexpected source. I published a parenthetical remark about the fox terrier issue (see Essay 11), ending with a serious point: “I also wonder what the textbook tradition of endless and thoughtless copying has done to retard the spread of original ideas.”
I have, over the years, maintained a correspondence about our favorite common subject with Roger Angell of the
New Yorker
, who is, among other things, the greatest baseball writer ever. I assumed that his letter of early April would be a scouting report for the beginning of a new season. But I found that Roger Angell is a man of even more dimensions than I had realized; he is also a fox terrier fancier. He had read my parenthetical comment and wrote, “I am filled with excitement and trepidation at the prospect of writing you a letter about science instead of baseball.”
Angell went on to suggest a fascinating and plausible explanation for the origin of the fox terrier simile (no excuse, of course, for its later cloning). Fox terriers were bred “to dig out foxes from their burrows, when a fox had gone to earth during a traditional British hunt.” Apparently, generations of fox-hunting gentlemen selected fox terriers not only for their functional role in the hunt but also under a breeder’s artifice to make them look as much like horses as possible. Angell continues, “The dogs rode up on the saddle during the hunt, and it was a pretty conceit for the owner-horseman to appear to put down a little simulacrum of a horse when the pack of hounds and the pink-coated throng had arrived at an earth where the animal was to do his work.” He also pointed out that fox terriers tend to develop varied patches of color on a basically white coat and that a “saddle” along the back is “considered desirable and handsome.” Thus, Angell proposed his solution: “Wouldn’t it seem possible that some early horse geologist, in casting about for the right size animal to fit his cliché-to-be, might have settled, quite unconsciously, on a breed of dog that fitted the specifications in looks as well as size?”
This interesting conjecture led me to devise the following, loosely controlled experiment. I asked David Backus, my research assistant, to record every simile for
Hyracotherium
that he could find in the secondary literature of texts and popular books during more than a century since O. C. Marsh first recognized this animal as a “dawn horse.” We would then use these patterns in attempting to locate original sources for favored similes in the primary literature of vertebrate paleontology. We consulted the books in my personal library as a sample, and compiled a total of eighty-six descriptions. The story turns out to be much more ascertainable and revealing than I had imagined.
The tradition of simile begins at the very beginning. Richard Owen, the great British anatomist and paleontologist, described the genus
Hyracotherium
in 1841. He did not recognize its relationship with horses (he considered this animal, as his chosen name implies, to be a possible relative of hyraxes, a small group of Afro-Asian mammals, the “coneys” of the Bible). In this original article, Owen likened his fossil to a hare in one passage and to something between a hog and a hyrax in another. Owen’s simile plays no role in later history because other traditions of comparison had been long established before scientists realized that Owen’s older discovery represented the same animal that Marsh later named
Eohippus
. (Hence, under the rules of taxonomy, Owen’s inappropriate and uneuphonious name takes unfortunate precedence over Marsh’s lovely
Eohippus
—see Essay 5 on the rules of naming.)
The modern story begins with Marsh’s description of the earliest horses in 1874. Marsh pressed “go” on the simile machine by writing, “This species was about as large as a fox.” He also described the larger descendant
Miohippus
as sheeplike in size.