Life on a Young Planet (12 page)
Read Life on a Young Planet Online
Authors: Andrew H. Knoll

At present, that’s about all we can say. The heat-scorched hills of North Pole suggest that life existed 3.5 billion years ago, and that, by itself, is remarkable. Warrawoona communities may have included photosynthetic microorganisms and other microbes with metabolisms still seen today. But many uncertainties persist. Warrawoona paleobiology remains a shadow play whose apparently familiar themes may be deceptive.
Northwestern Australia is one of two places in the world that contain well-preserved sedimentary rocks as old as 3.5 billion years. The other is the rugged Barberton Mountain Land, near Kruger Park in South Africa. The two areas are similar, so much so that some geologists believe that they form parts of a single ancient terrain, severed by plate tectonic
movements long after the Archean. Their paleontological inventories bear comparison, as well. Both include stromatolites of uncertain origin, along with organic matter whose carbon isotopic composition and sedimentary distribution suggest some type of photosynthesis. Both have been heated to temperatures that destroy biomarker molecules. And like those of Warrawoona, Barberton cherts contain spherical and filamentous microstructures reminiscent of fossils.
As a graduate student, I had a go at Archean paleontology, traveling to Africa as Elso Barghoorn’s field assistant. Having grown up on Tarzan books, I was excited as the plane touched down in Johannesburg late at night. I couldn’t wait to catch my first glimpse of Africa the next morning, and was only a little disappointed that the view from my hotel window looked a lot like Chicago. Within hours, we were on the road, and as the cityscape receded in the rearview mirror, the great South African veldt opened before us. Culturally, ecologically, and geologically, the Barberton Mountain Land was new to me. In each clump of thorn trees I sensed menace, and in each chert I espied fame. Neither fame nor menace materialized, but the cherts did turn out to contain microstructures that are probably, if not unambiguously, biological.
In one particular sample marked by centimeter-scale stromatolite-like precipitates, I discovered a large population of spherical microstructures 2 to 4 microns in diameter—the size and shape of small cyanobacteria (
figure 4.6a
). The structures occur in individual laminae. Moreover, they are made of organic matter, and some preserve both an outer wall and a raisinlike interior body, also organic. The microstructures are compressed along the bedding surface, much like younger microfossils—indeed, this slight flattening tells us that the structures formed before enclosing sediments were compacted by burial. The distribution of sizes in the population matches that of modern cyanobacteria, and the structures also show evidence of binary division, again much like living blue-greens.
So are these fossil cyanobacteria? Not necessarily. Many different bacteria are small and spherical. More sobering, nonbiological processes can, in principle, produce similar structures—although it isn’t obvious that such processes were at work in the Barberton seaway. The Barberton spheres, thus, fall only a few steps ahead of Warrawoona filaments. They could be fossil cyanobacteria or some other type of microorganism. They
could record a primordial microbe that is long extinct. Or they could be carbonaceous spheres formed by physical processes on the Barberton seafloor. We simply don’t know. More recently, Maud Walsh of Louisiana State University has made a careful study of the organic matter in Barberton cherts, finding bedding textures most easily explained as mats and thin filaments that may be microfossils.
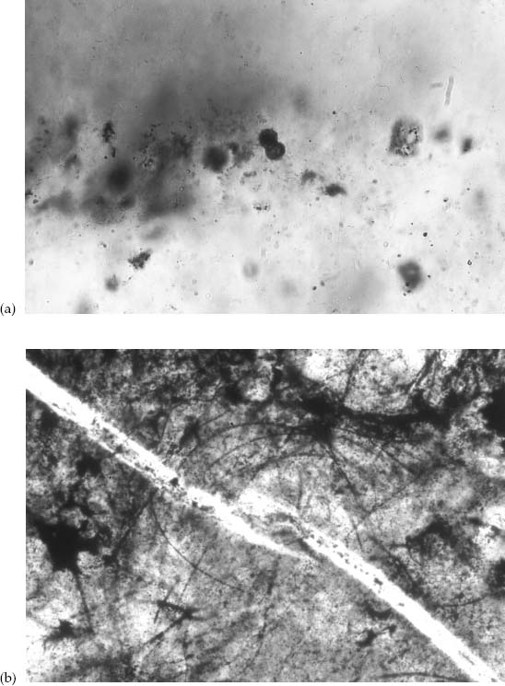
Figure 4.6.
(a) Carbonaceous microstructure, possibly preserving a microbe during cell division, in 3.4-billion-year-old rocks from South Africa. Sphere is 4 microns in diameter. (b) Filamentous microfossils in 3.2-billion-year-old rocks from northwestern Australia. Each filament is about 2 microns across. (Photo (b) courtesy of Birger Rasmussen)
What kind of planet can we piece together from these fragments? Geologically, it appears to have been a world of familiar processes but not-so-familiar patterns. Continents began to form at least 4.2 billion years ago, and chemical details of volcanic rocks from Barberton, Warrawoona, and other old terrains suggest that a large volume of continental crust had formed by the time they were deposited. Little of these early continents remains, however, implying that on the early Earth, continents were recycled back into the mantle more easily than they are today. Three and a half billion years ago, plate tectonics had already begun to pattern our planetary surface, but Earth’s upper mantle appears to have been hotter, the basaltic crust beneath the oceans thicker, and, perhaps, the continents smaller and less stable. Then, as now, continental crust probably formed at plate margins, where descending slabs of oceanic crust cause overlying rocks to melt. On the other hand, early continent formation may have received a significant boost from a source that is no longer important—partial melting of basalts buried beneath thick piles of lava spilled onto the seafloor.
The rock record that survives from the early Earth is not simply the fragment of a geologically modern planet buffeted by time. Something about the character and mix of processes that form and destroy continents was different, and though many insightful scientists have hazarded opinions, we don’t fully understand what it was.
We have a bit more confidence that when the Warrawoona seaway formed, Earth was a biological planet. Moreover, the evidence of carbon isotopes suggests that the great ecological liberation of photosynthesis may already have begun. Whether or not contemporary microorganisms included the oxygen-producing cyanobacteria is uncertain, but the presence of
any
type of photosynthetic organism in the Warrawoona ocean speaks volumes, because it allows us to place a calibration point on the Tree of Life introduced in
chapter 2
. In the new view of microbial
evolution symbolized by the tree, photosynthetic organisms are relative latecomers that diversified long after the origin of life and the divergence of biology’s principal domains. If Warrawoona organic matter was made by photosynthesis, then a great deal of evolution must already have taken place.
Microorganisms appear to have cycled carbon, sulfur, and nitrogen through early Archean ecosystems, just as they do today. There is no record of eukaryotes or archaeans in these oldest rocks, but then there aren’t many fossils, period, and it would be hazardous to interpret the absence of evidence as evidence for absence. The branching pattern of the Tree of Life tells us that if photosynthetic bacteria lived in the Warrawoona sea, then at least some Archaea were almost certainly present.
We have one more constraint on early Archean biology. Consistent with our environmental reading of the tree, geological observations indicate that 3.5 billion years ago Earth’s atmosphere contained nitrogen, carbon dioxide, and water vapor but little free oxygen. Most inferences about ancient environments are gleaned from subtle geochemical clues, but the sedimentary signature of oxygen limitation is flamboyant—bright red bands of cherty rock rich in the iron oxide mineral hematite (Fe
2
O
3
). Aptly named banded iron formation (BIF, for short), these rocks do not form in the present-day ocean. In fact, with one important exception, they haven’t accumulated for the past 1.85 billion years. But for the first half of Earth history, BIFs were a standard component of marine sediments. The reason that BIFs do not form today is that iron entering the oceans immediately encounters oxygen and precipitates as iron oxide; as a result, iron concentrations in modern seawater are extremely low. The BIF in Archean sedimentary successions may have formed by the reaction of iron with oxygen. Or the iron may have been oxidized by photosynthetic bacteria or UV radiation that penetrated to the sea surface in the absence of an effective ozone shield. However they were precipitated, the BIFs tell us that on the early Earth, iron didn’t get stripped away as it entered the sea. Instead, iron was readily transported in solution throughout the deep ocean. This could only happen if deep waters were free of oxygen, forcing us to conclude that the Archean atmosphere and the sea surface in contact with that atmosphere had much less oxygen than today. Just how much oxygen was present is contentious, but it couldn’t have been more than about 1 percent of presentday
levels and may have been much less. Under these conditions, aerobic respiration and chemosynthetic metabolisms that depend on molecular oxygen may have been limited or absent, depending on just how much oxygen was available.
Together, then, geology and microbiology suggest that early Archean oceans differed from those of younger intervals, containing much less oxygen but more iron. The early ocean may have been warmer than today’s, but geology provides us with few real constraints on Archean climate. All we can say is that if photosynthesis was present, surface waters could have been no warmer than 74°C—the maximum temperature tolerated by photosynthetic organisms. The Tree of Life suggests that the earliest Bacteria and Archaea lived at higher temperatures, but this need not be interpreted to mean that the entire ocean was hot. Early heat-loving microbes might well have lived in hydrothermal environments like those inhabited by their descendants today.
How can we amplify the weak biological signal in early Archean rocks? Two strategies come readily to mind; one concerns how we look for evidence and the other, where we search. As the saga of Warrawoona illustrates, the paleontological search strategy that has worked so well in younger rocks—collect black chert, and lots of it—has not been notably successful in early Archean rocks. Malcolm Walter, a leading Australian paleontologist, has advocated a different kind of search, one that focuses on the very hydrothermal processes that ravaged the contents of preserved cherts. As noted earlier, hydrothermal systems are home to some of the earliest branching organisms on the Tree of Life; moreover, hydrothermal springs commonly deposit carbonate and silica minerals that might preserve records of early biology. Chimneys of pyrite and other minerals also form at some hydrothermal vents in the ocean. Comparable deposits occur in early Archean terrains, but until recently they have not received much attention from paleontologists. This situation is changing, however, and early in 2000, Australian geologist Birger Rasmussen reported convincingly biological (if metabolically uninformative) filaments from hydrothermal mineral deposits that, at 3.2 billion years, are not much younger than Warrawoona (
figure 4.6b
). I anticipate that as our search strategy widens, our understanding of early biology will grow proportionately.
The second obvious research path is to discover older rocks. Because of our planet’s restless surface, metamorphism, uplift, and erosion continually alter and destroy the rock record—successively older periods of time are represented by ever smaller volumes of rock. This being the case, it is no simple task to find little-altered sedimentary rocks older than the Warrawoona and Barberton successions. Yet, Roger Buick has done just that. In a remote area of northwestern Australia, he discovered a succession of sedimentary and volcanic rocks that lies
beneath
the Warrawoona Group. Christened the Coonterunah succession, these beds contain volcanic rocks dated at 3,515 ± 3 million years—not dramatically older than the Warrawoona rocks that cover them, but nonetheless a step deeper into Earth history. Coonterunah rocks include sedimentary beds deposited in deepwater environments along with basaltic lavas, but to date they have not yielded fossils.
Steve Mojzsis and his colleagues took a bigger step backward—to cirea 3.8-billion-year-old rocks on Akilia Island off the coast of southwestern Greenland. These rocks have been severely altered by metamorphism, making it difficult to read their geologic history. Mojzsis and colleagues interpret the rocks as sediments laid down on an ancient seafloor. Within the rocks, they found tiny grains of mineral phosphate, and, within these mineral grains, still tinier inclusions of reduced carbon (graphite). Using an ion microprobe, Mojzsis measured the isotopic composition of the carbon and found that it is strongly depleted in
13
C, consistent with formation by biological processes.
But nothing is ever simple in Archean geology. After careful reexamination of the Akilia rocks, a team led by phosphate expert Gus Arrhenius (one of the “and colleagues” in Mojzsis’s original group) concluded that the phosphate grains formed relatively late in the history of these rocks, during alteration by hot metamorphic fluids. Moreover, Arrhenius and colleagues believe that the graphite in these grains formed at the same time, by chemical reaction of metamorphic fluids with iron carbonate in the rock. Independent research by geologists Christopher Fedo and Martin Whitehouse supports a metamorphic origin for key features of the Akilia rocks. Indeed, Fedo and Whitehouse believe that the rocks originated as igneous bodies deep within the Earth’s interior.
Here’s one more debate still to be resolved. But if the revised interpretation is correct, the carbon in Akilia rocks can tell us nothing about
life. In fact, Arrhenius’s conclusions undermine more than that. Recall that those graphite crystals in Akilia phosphate grains have carbon-isotope ratios much like organic matter produced by photosynthesis. If physical processes can fractionate carbon isotopes by up to 50 parts per thousand, then our faith in carbon-isotope composition as a biosignature is shaken.