Neanderthal Man (3 page)

By this time, I was all too acquainted with the fact of contamination. I had been working for more than twelve years on the extraction and analysis of ancient DNA from such extinct mammals as cave bears, woolly mammoths, and ground sloths. After a string of frustrating results (I detected human mtDNA in almost all the animal bones I analyzed with the PCR), I spent a great deal of time thinking about, and devising, ways to minimize contamination. Thus, Matthias performed all extractions and other experiments, right up to the first temperature cycle of the PCR, in a small lab that was kept meticulously clean and absolutely separate from the rest of our laboratory. Once the ancient DNA, the primers, and the other components necessary for the PCR had been placed together in a tube, the tube was sealed, and the temperature cycles and all subsequent experiments were performed in the regular laboratory. In the clean lab, all surfaces were washed with bleach once a week, and every night the lab was irradiated with UV light to destroy any dust-borne DNA. Matthias entered the clean lab through an antechamber, where he and others working there donned protective gowns, face shields, hairnets, and sterile gloves. All reagents and instruments were delivered directly to the clean lab; nothing was allowed into it from other parts of the institute. Matthias and his colleagues had to start their workday in the clean lab instead of in other parts of our laboratory, where large amounts of DNA were being analyzed. Once they had entered any such lab, they were banned from the clean lab for the rest of the day. To put it mildly, I was paranoid about DNA contamination, and I felt I had good reason to be.
Even so, in Matthias’s initial experiments we had seen evidence of some human contamination of the bone. After using the PCR to amplify a piece of mtDNA from the bone, he had cloned the resulting batch of supposedly identical DNA copies in bacteria. He did this in order to see whether more than one type of mtDNA sequence might exist among the cloned molecules: each bacterium will take up just one 61-nucleotide molecule joined to a carrier molecule called a plasmid and grow up to a clone of millions of bacteria where each carries copies of the 61-nucleotide molecule the first bacterium took up, so we were able to get an overview of whatever different DNA sequences existed in the population of molecules by sequencing a number of the clones. In Matthias’s earliest experiment, we saw seventeen cloned molecules that were similar or identical to one another while different from the two thousand–plus modern human mtDNAs we were using for comparison—but we also saw one that was identical to a sequence seen in some humans today. This clearly showed the presence of contamination, perhaps from museum curators or others who had handled the bone over the 140 years since its discovery.
So the first thing Matthias did in his attempt to reproduce our original result was to repeat the PCR and the cloning. This time he found ten clones with the unique sequence that had excited us so much and two that looked like they could have come from any modern person. He then went back to the bone and made another extract, did the PCR and cloning again, and got ten clones of the interesting type and four that looked like mtDNA from present-day humans. Now we were satisfied: our original result had passed the first tests—we could repeat them and see the same unusual DNA sequence each time.
Matthias next started to “walk along” the mtDNA, using other primers designed to amplify fragments that overlapped a part of the first fragment but extended further into other regions of the mtDNA (see Figure 1.2). Once again, we observed that some of the sequences of these fragments had nucleotide changes never seen in contemporary humans. Over the next few months, Matthias amplified thirteen different DNA fragments of different sizes, each at least twice. The interpretation of the sequences was complicated by the fact that any one DNA molecule can carry mutations that can be due to various causes: ancient chemical modifications, sequencing errors, or even rare but natural variation that may exist among the mtDNA molecules found within a cell of an individual. Therefore, we used a strategy I had worked out previously for ancient animal DNA (again, see Figure 1.2). For each position in each experiment, we took as authentic the so-called consensus nucleotide—that is, the nucleotide (A, T, G, or C) carried in that position by most of the molecules we examined. We also required that each position be identical in at least two independent experiments, since a PCR might, in an extreme case, have started from a single DNA strand—in which case all the clones, due to some error in the first PCR cycle or to some chemical modification in that particular DNA strand, would carry the same nucleotide in the same position. If two PCRs differed with regard to even a single position, we repeated the PCR a third time, to see which of the two nucleotides was reproducible. Matthias eventually used 123 cloned DNA molecules to puzzle together a sequence of 379 nucleotides of the most variable part of the mtDNA. According to the criteria we had settled on, this was the DNA sequence that this Neanderthal carried when he or she was alive. Once we had this longer sequence, we could begin the exciting work of comparing it with the variations seen in present-day humans.

Figure 1.2. Reconstruction of a piece of mtDNA from the Neanderthal from Neander Valley. Above, a modern reference sequence is shown. Each line below represents one cloned molecule amplified from the Neanderthal type specimen. Where these sequences are identical to the reference sequence, I have placed a dot; where they differ from the nucleotide, I have written them out. In the bottom line is the reconstructed Neanderthal nucleotide sequence. At each position, we require that a change from the reference sequence is seen in a majority of clones and in at least two independent PCR experiments (either the ones shown or others). From Matthias Krings et al., “Neandertal DNA sequences and the origin of modern humans,”
Cell
90, 19–30 (1997).
At this point, we compared our 379-nucleotide Neanderthal mtDNA sequence to the corresponding mtDNA sequences from 2,051 present-day humans from all around the world. On average, twenty-eight of the positions differed between the Neanderthal and a contemporary person, whereas people alive today carry an average of only seven differences from one another. The Neanderthal mtDNA was four times as different.
We then looked for any indication that the Neanderthal mtDNA was more like the mtDNA found in modern Europeans. One might well expect to find this, since Neanderthals evolved and lived in Europe and western Asia; indeed, some paleontologists believe that Neanderthals are among the ancestors of today’s Europeans. We compared the Neanderthal mtDNA with that of 510 Europeans and discovered that it carried, on average, twenty-eight differences. We then compared it with mtDNA from 478 Africans and 494 Asians. The average number of differences from the mtDNAs of these people was also twenty-eight. This meant that, on average, European mtDNAs were no more similar to the Neanderthal mtDNA than were mtDNAs from modern-day Africans and Asians. But maybe Neanderthal mtDNAs were similar to mtDNAs found in just some Europeans, as one would expect if Neanderthals had contributed some mtDNA to Europeans. We checked this and found that the Europeans in our sample whose mtDNAs were most like that of the Neanderthal showed twenty-three differences; the Africans and Asians closest to the Neanderthal carried twenty-two and twenty-three, respectively. In short, we observed not only that the Neanderthal mtDNA seemed very different from the mtDNAs of modern humans worldwide but also that there was no indication of any special relationship between the Neanderthal mtDNA and any subset of European mtDNAs alive today.
However, just counting differences is not enough to reconstruct the evolutionary history of a piece of DNA. The differences found between DNA sequences represent mutations that occurred in the past. But some types of mutations are more frequent than others, and some positions in DNA sequences are more prone to mutate than others. At such positions, more than one mutation—especially the types that happen more frequently—may have occurred in the history of a DNA sequence. Therefore, to estimate the history of this particular piece of mtDNA, we needed to apply models for how we believed it had mutated and evolved, bearing in mind that certain positions might have mutated more than once, thus obscuring previous mutations. The result of such a reconstruction is depicted as a tree, in which a DNA sequence on the tip of a branch links back to a common ancestral DNA sequence. These ancestral sequences are depicted as the points where branches join on the tree (see Figure 1.3). When we did such a tree reconstruction, we saw that the mtDNAs of all humans alive today trace their ancestry back to one common mtDNA ancestor.
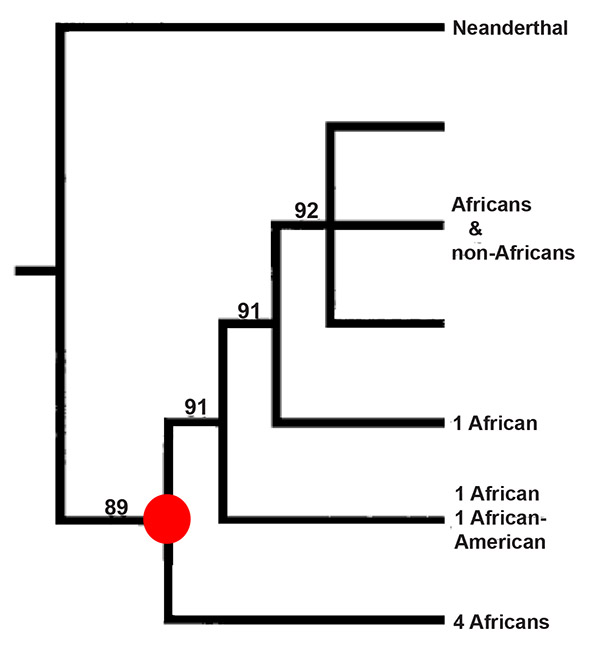
Figure 1.3. A mtDNA tree, illustrating how the mtDNAs of people alive today trace their ancestry back to a common mtDNA ancestor (the so-called Mitochondrial Eve, indicated by a circle) who existed more recently than the mtDNA ancestor shared with the Neanderthal. Nucleotide differences are used to infer branching order, and the numbers refer to the statistical support for the branching order shown. Modified from Matthias Krings et al., “Neandertal DNA sequences and the origin of modern humans,”
Cell
90, 19–30 (1997).
This finding, which was already known from Allan Wilson’s work in the 1980s,
{1}
is in fact expected for mtDNA, since each of us carries only a single type and cannot exchange pieces of it with other mtDNA molecules in the population. Since mtDNA is passed on only by mothers, the mtDNA lineage of a woman will die out if she has no female descendants—so in each generation some mtDNA lineages vanish. Therefore, there must once have existed a woman—the so-called Mitochondrial Eve—who carried an mtDNA lineage that would turn out to be the ancestor of all human mtDNAs today, simply because all other lineages since that time have been lost, purely by chance.
According to our models, however, the Neanderthal mtDNA did not trace back to this Mitochondrial Eve but went further back before it shared an ancestor with the mtDNAs of humans alive today. This finding was immensely exciting. It proved beyond any doubt that we had recovered a piece of Neanderthal DNA—and it showed, at least with respect to their mtDNA, that the Neanderthals were profoundly different from us.
My colleagues and I also used the models to estimate how long ago the Neanderthal mtDNA shared an ancestor with current human mtDNAs. The number of differences between the two types of mtDNA is an indication of how long they have been transmitted through generations independently of each other. The mutation rates of widely separated species—mice and monkeys, say—will differ, but among closely related species—such as humans, Neanderthals, and the great apes—they have been constant enough to allow scientists to estimate, based on the differences observed, when two DNA sequences last shared an ancestor. Using the models for how fast different types of mutations occur in mtDNA, we estimated that the mtDNA ancestor common to all humans alive today, the Mitochondrial Eve, lived between 100,000 and 200,000 years ago, as Allan Wilson and his team had found. However, the ancestor that the Neanderthal mtDNA shared with human mtDNAs lived about 500,000 years ago; that is, she was three or four times as ancient as the Mitochondrial Eve from whom all present-day human mtDNAs are descended.
This was amazing stuff. I was now fully convinced that we had recovered Neanderthal DNA and that it was very different from the DNA of modern humans. However, before publishing our findings, we needed to overcome the last hurdle: we needed to find an independent laboratory that could repeat what we had done. Such a lab would not need to determine the entire 379-nucleotide mtDNA sequence, but it
would
have to retrieve one of the regions that carried one or more substitutions setting Neanderthals apart from humans today. This would show that the DNA sequence we had determined really existed in the bone and wasn’t some strange and unknown sequence perhaps floating around in our laboratory. But to whom could we turn? This was a delicate issue.