The Cerebellum: Brain for an Implicit Self (27 page)
Read The Cerebellum: Brain for an Implicit Self Online
Authors: Masao Ito
Tags: #Science, #Life Sciences, #Medical, #Biology, #Neurology, #Neuroscience

A unique method of testing for cerebellar-evoked adaptability during locomotion was introduced by the use of split-belt treadmill locomotion in which the speed of each belt (i.e., for the right- versus left-side legs) could be controlled independently. While decerebrate cats were performing controlled locomotion on such a device, both fore- and hindlimbs on one side were suddenly subjected to a higher belt velocity. The cat gradually adapted using an unusual (atypical) pattern of limb coordination to retain stable locomotion. An inhibitor of NO synthase or a scavenger of NO applied to the cerebellar locomotion area abolished this adaptation. Because NO is required for the
induction of conjunctive LTD, this finding supported the hypothesis that learning in locomotion was based on the induction of conjunctive LTD (
Yanagihara and Kondo, 1996
). The adaptive behavior described here has been well reproduced by simulation using an autonomous distributed control model of locomotion (
Ito et al., 1999
).
In human patients with cerebellar lesions, two types of gait abnormality were categorized: one featured by impaired leg placement and the other by deficiencies in dynamic balance control. The former abnormality included inadequate adaptation to added weight loads. The cerebellar lesions in patients with the former deficiencies usually involved the interpositus and adjacent dentate nuclei. In contrast, patients with the latter abnormality usually had lesions in the fastigial nuclei (and to a lesser degree the interpositus nuclei) (
Ilg et al., 2008
).
Split-belt treadmill locomotion has also been applied to humans to study their interlimb coordination (Diezt et al., 1994). In Morton and Bastian’s paradigm, belt speed was switched in sequential steps from both-slow (0.5 meters/second), to both-fast (1 meter/second), to both-slow, to the split-belt condition (one fast, the other slow), and finally to both-slow. Both normal control subjects and cerebellar patients were able to quickly adjust their stride length (distance traveled by one of the ankle markers from the time of initial contact to the time of lift-off of one limb) and percentage length of the stance phase (stride length in percentage of the total time from initial contact to the next initial contact). In contrast, the step length (difference between the distances of the leading and trailing limb ankle markers at the time of initial contact on the leading limb) and the double limb support time (time from initial contact on one limb to lift-off of the other limb, expressed as a percentage of the total stride time of the lift-off limb) changed slowly in the control subjects, and these adaptations to the split-belt condition could not be achieved in the cerebellar patients. Hence, it appeared that the former two parameters were determined by reactive feedback-driven adaptation and the latter two parameters by predictive feedforward adaptation in the cerebellum (
Morton and Bastian, 2006
).
The classical conditioning of the eye-blink reflex, particularly “delay conditioning” in which conditioned and unconditioned stimuli are contiguous, has been introduced as a simple and robust model of cerebellar learning (
Lincoln et al., 1982
;
Yeo et al., 1985a
,
b
,
c
). Conditioned eye-blink responses develop during repeated trials. Once established, they remain (consolidation), unless they are extinguished by unpaired conditioned and unconditioned stimuli.
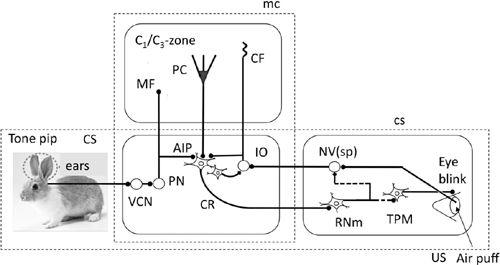
The conditioned reflex pathway (
Figure 35
) is mediated by a group of neurons in the anterior interpositus nucleus. These neurons in the rabbit receive conditioned
stimuli such as a tone pip and, in turn, send signals via the red nucleus to trigeminal premotor and motor neurons. These latter cells induce eyelid closure, eyeball retraction, and a resulting passive extension of the nictitating membrane. Anterior interpositus neurons are normally inhibited by Purkinje cells in lobule HVI, which are activated by mossy fiber-mediated tone-pip signals. When, however, unconditioned stimuli, such as an air puff, were applied to the cornea to evoke climbing fiber signals, conjunctive LTD released the anterior interpositus neurons from Purkinje cell inhibition. This was shown to occur at the precise moment that the air puff was applied (
Jirenhed et al., 2007
; see also McCormick and Thompson, 1984; Berthier and Moore, 1986). Thus, an eye blink was evoked in a timely manner to protect the cornea from the air puff. Later studies revealed that impaired eye-blink conditioning occurred in mutant mice that lacked conjunctive LTD (
Shibuki et al., 1996
;
Kishimoto et al., 2001
,
2002
;
Miyata et al., 2001
).
Figure 35. Wiring diagram for eye-blink conditioning.
Tone pip-evoked auditory signals as the conditioned stimulus (CS) enters the cerebellum via projections from the pontine nuclei (PN). Air puff as the unconditioned stimulus (US) enters the cerebellum via projections from the inferior olive (IO). The CS and US pathways converge onto the nucleus interpositus (AIP). The postsynaptic conditioned response CR pathway projects from the AIP to the magnocellular red nucleus (RNm) and ultimately connects to neurons (TPM) that cause eye blinking. TPM includes trigeminal premotor neurons, motoneurons for the eyelid muscles and, in rabbit, the nictitating membrane. Additional abbreviations: CS, broken line square enclosing control system structure; MC, that for microcomplex; NV(sp), spinal trigeminal nucleus; VCN, ventral cochlear nucleus. Symbols: broken line, polysynaptic connections. Other conventions are similar to
Figure 33
. (Based on
Thompson, 1988
.)
“Blocking” studies of eye-blink conditioning provide the opportunity to define the roles of the nucleo-olivary inhibitory projection (N-O in Color Plate V).
Recordings of cerebellar neuronal activity have shown that the inputs of the IO to the cerebellum become suppressed as learning occurs. This corresponds to the behavioral phenomenon of “blocking.” It occurs when a conditioned stimulus (CSA) is first paired extensively with an unconditioned stimulus (US), and then a second conditioned stimulus (CSB) is combined with CSA and the same US. Virtually no conditioning then occurs for the CSB. This blocking effect suggests that if an US is already fully predicted by one stimulus, and if the addition of a new stimulus provides no new information about the US, then the US will not activate or support the learning process responsible for establishing a new CS-US association. Because disrupting the inhibition of the IO by local infusion of a GABA antagonist, picrotoxin, prevented blocking in rabbits, the nucleo-olivary negative feedback process could be the neural mechanism mediating blocking (
Kim et al., 1998
). The disruption of nucleo-olivary inhibition also prevented the extinction of eye-blink conditioning, whereas that of excitatory inputs induced extinction (
Medina et al., 2002
). These findings are interpreted as indicating that the nucleo-olivary inhibition is required for setting the climbing fiber discharge rate at a level appropriate for driving extinction. However, caution is needed in interpreting these observations because the manipulation of olivary neuronal activity influences the background activity of Purkinje cells. This might affect learning and its behavioral manifestation (
Bengtsson et al., 2004
).
The effects of the removal of the cerebellar cortex and the local application of pharmacological agents on eye-blink conditioning have been controversial in studies carried out to determine whether memory is stored in the cerebellar cortex or alternatively in the anterior interpositus nucleus (for review, see
Christian and Thompson, 2003
). The importance of the cerebellar cortex was emphasized because the acquisition training for eye-blink conditioning was blocked when the anterior lobe of the cerebellar cortex was lesioned (
Garcia et al., 1999
) or when the lobule HVI was inactivated by localized infusion with an AMPA/kainate receptor blocker, CNQX (
Attwell et al., 2001
). A specific contribution of the cerebellar cortex to the timing of eye-blink conditioning was suggested because a reversible blockade of cerebellar cortical outputs via the infusion of picrotoxin into the interpositus nucleus disrupted the timing to accurately coincide conditioned responses with the unconditional stimulus. In the same animals, conditioned responses were abolished by the infusion of a GABA agonist, muscimol (
Garcia and Mauk, 1998
; Bao et al., 2002). In mice lacking conjunctive LTD owing to the Purkinje cell-specific inactivation of PKC, conditioned responses substantially remained, but accurate timing was lost (
Koekkoek et al., 2003
).
The abnormally timed, short-latency conditioned responses remaining after the blockade of Purkinje cell outputs could be generated by an enhanced excitation of anterior interpositus neurons. This postulate was adopted in simulating eye-blink conditioning (
Medina et al., 2000
). Four lines of experimental evidence supported this idea. (1) Anterior interpositus nuclear neurons displayed an associative form of learning plasticity (
Ohyama et al., 2006
). (2) Synapse formation occurred in the interpositus nucleus in association with eye-blink conditioning (
Kleim et al., 2002
). (3) The injection of a protein synthesis inhibitor, anisomycin, in the bilateral cerebellar hemispheres reversibly blocked the acquisition, even though it did not block the performance of a conditioned response (
Bracha et al., 1998
). (4) Eye-blink-conditioned animals exhibited a new category of cell-division-cycle-2 (cdc2)-related kinases in interpositus neuron mRNAs (
Gomi et al., 1999
).
A more recent study showed that consolidation was prevented by the application of the GABA agonist, muscimol, shortly after training had inactivated the cerebellar cortex. Post-training inactivation of the interpositus nucleus, in contrast, allowed conditioning to develop normally (
Attwell et al., 2002
). These observations indicate that a cortical circuit is essential for memory consolidation. A complication arises, however, from the possibility that memory consolidation is cortical in the cerebellum but sensitive to the increased excitability of nuclear neurons following the loss of cortical inhibition. When cerebellar lobule HVI and the anterior interpositus nucleus of rabbits were inactivated simultaneously during the post-training period, consolidation was still impaired. This suggests that the disinhibitory effects of cortical inactivation on the interpositus nucleus are irrelevant for consolidation (
Kellett et al., 2010
).
A crucial role of conjunctive LTD in eye-blink conditioning has been suggested in two ways. Activation of PKC, which is required for conjunctive LTD induction (
Chapter 7
, “
Conjunctive Long-Term Depression (LTD)
”), occurred within the molecular layer of lobule HVI in rabbits given this conditioning (
Freeman et al., 1998
). On the other hand, mice lacking conjunctive LTD owing to the Purkinje cell-specific inactivation of PKC retained their conditioned responses, but its accurate timing was lost (
Koekkoek et al., 2003
). Note, however, that the relationship of eye-blink conditioning to conjunctive LTD is not so simple because a mutant mouse lacking the fragile X mental retardation type 1 protein (FMR1) exhibited impaired eye-blink conditioning in its timing when they exhibited unusually enhanced LTD (
Koekkoek et al., 2005
). When the HVI areas of the rabbit cerebellum were sliced 24 hours after the rabbit had been trained for eye-blink conditioning, sequentially applied parallel fiber and climbing fiber stimuli failed to induce conjunctive LTD, whereas it occurred in cerebellar slices dissected from control rabbits (
Schreurs et al., 1997
). This suggests that conjunctive LTD underlying eye-blink conditioning continues for at least 24 hours and occludes the elicitation of another conjunctive LTD. An increased excitability of Purkinje cell dendrites in
the rabbit lobule HVI was detected and sustained for one month following the initial acquisition of eye-blink conditioning (Schreurs et al., 1998). This learning-related excitability increase is presumably caused by changes in K
+
current, possibly mediating an I
A
-like current, but its relationship to conjunctive LTD is unclear. Finally, a significant decrease in the number of excitatory synapses in the molecular layer of the cerebellar cortex has been reported in eye-blink-conditioned rabbits (
Connor et al., 2009
).
Whereas evidence continues to accumulate for cerebellar plasticity playing a key role in eye-blink conditioning, a recent study by Nakanishi’s group suggested a more complex possibility. This study used reversible neurotransmission blocking (RNB) mice (Wada et al., 2007). In animals treated this way, only granule cells expressed tetanus toxin in a doxycycline (DOX)-dependent manner. Simple spikes of Purkinje cells elicited by granule cell inputs were turned off and on by DOX treatment and its withdrawal, respectively. It was found that blockade of granule cell inputs to Purkinje cells abolished eye-blink conditioned responses in a DOX-dependent manner. Thus, a cortical circuit via Purkinje cells appeared to be essential for expressing the conditioned responses. Peculiarly, however, from the beginning of the reconditioning process, when granule cell inputs recovered by removal of DOX, seemingly normal conditioned responses were immediately produced in the reconditioned mice. Bilateral destruction of the interpositus nuclei prior to eye-blink conditioning abolished the learning. These results can be explained on the basis of a bilateral membrane-potential dependent synaptic plasticity at mossy fiber-nuclear neuron synapses (depressed by depolarization and potentiated by hyperpolarization), as recently reported by McElvain et al. (
2010
). It might happen that mossy fiber synapses in the interpositus nucleus are depressed by the depolarization caused by DOX turning off of Purkinje cell signals. The effect might then be reversed to potentiation quickly when membrane hyperpolarization is caused by reactivated Purkinje cells. The potentiated conditioned responses might look like learned responses.