The Cerebellum: Brain for an Implicit Self (30 page)
Read The Cerebellum: Brain for an Implicit Self Online
Authors: Masao Ito
Tags: #Science, #Life Sciences, #Medical, #Biology, #Neurology, #Neuroscience

For the withdrawal reflex of a forelimb, the anterior interpositus nucleus acts as a controller, whereas the magnocellular red nucleus neurons, forelimb flexor motoneurons, and associated segmental neurons are nested as a controlled object (
Figure 33
). A microcomplex involving the C
1
/C
3
-zone and the anterior interpositus nucleus constitutes an adaptive controller.
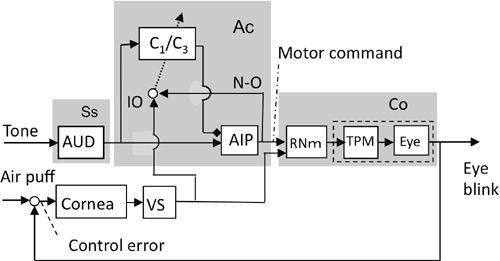
The unconditioned eye-blink reflex is evoked by an air puff, which drives trigeminal premotor neurons and motoneurons and induces eyelid closure. The magnocellular red nucleus forms a long loop pathway superposed on the eye-blink reflex pathway. Based on the neuronal circuit so far dissected (
Figure 35
), the block diagram in
Figure 41
is formulated. There, the anterior interpositus nucleus, together with the HVI area of the C
1
- and C
3
-zones, form an adaptive controller of the eye-blink reflex, whereas the magnocellular red nucleus is nested to the controlled object together with trigeminal premotor neurons and motoneurons. Normally, tone pip stimuli do not drive the eye-blink reflex, but after classic eye-blink conditioning, they evoke an eye blink in a timely and predictive manner in order to protect the cornea from the air puff. This adaptive mechanism is now explained by a combination of conjunctive LTD that is induced by tone-pip-induced simple spikes, and air-puff-induced climbing fiber spikes in Purkinje cells. Another form of learning is also involved. It occurs in those synapses supplied by the tone-pipe-mediating mossy fibers to the anterior interpositus nuclear neurons (see
Chapter 9
,
Section 7
and
Chapter 11
,
Section 5
).
Figure 41. Control system scheme for eye-blink conditioning.
This block diagram is based on the wiring diagram illustrated in
Figure 35
. The cerebellum provides an adaptive controller for the controlled object, which in this case is the eye-blinking system including the magnocellular red nucleus (RNm). Whereas tone stimuli provide mossy fiber inputs, air puff stimuli provide climbing fiber inputs. Abbreviations: Ac, adaptive controller; AIP, anterior interpositus nucleus; AUD, auditory system; C1/C3, zones of lobules VI-VII responsible for eye-blink conditioning; Co, controlled object; IO, inferior olive; N-O, a nucleo-olivary inhibitory projection, which is presumed to be responsible for the “blocking” phenomenon (see text); TPM, trigeminal premotor neuron and motoneuron; VS, visual system. (Based on
Ito, 2006
.)
When the wiring and block diagrams for various reflexes are compared, two prototypes of microcomplexes stand out. For the VOR, OKR, OFR (
Figures 28
,
36
–
38
) and myotatic reflex (
Figure 32
), the controller is served by certain vestibular nuclear neurons, and hence the microcomplex has been called “V-type.” In contrast, for the limb withdrawal reflex (
Figure 33
), saccadic eye movement (
Figure 40
), and eye-blink conditioning (
Figure 41
), the controller is served by cerebellar nuclear neurons (fastigial or anterior interpositus), and the microcomplex involved has been called “C-type” (
Ito, 1984
). The V-type prevails in the flocculonodular lobe and B-zone (
Figures 28
,
32
), whereas the C-type is found in the A- and C
1
/C
3
-zones (
Figures 33
,
40
,
41
). These two types of microcomplexes may function similarly to each other as an adaptive controller, but they act on the controlled objects of different structures; V-types act on one-step-nested controlled objects, whereas C-types act on two-step-nested controlled objects (see
Figures 11
and
12
). We may consider that the C-type is more advanced than the V-type in an evolutionary sense in view of the higher integration level of the C-types’ controlled objects.
The cerebellum can now be viewed as an assembly of numerous microcomplexes, many of which are associated with reflexes to afford them a substantial degree of adaptability. This feature somewhat resembles the recently developed “embedded computing system,” in which a tiny computer is lavishly used in each instrument, such as a portable phone, in contrast to a large computer commonly shared by many users. Much is yet to be learned about the specific neuronal processes that operate in each microcomplex.
We now proceed to the examination of neuronal circuits for another category of movements: the voluntary control of arms, hands, and fingers, as commanded by the primary motor cortex. Importantly, this control involves the cerebrocerebellar communication loop that links the primary motor cortex and the intermediate part of the cerebellar hemisphere. This system enables humans to acquire skills for multijointed limb movements with many degrees of freedoms (27 for an arm with a hand and five fingers). Various test paradigms have been designed to study this form of cerebellar control. They include noting movement deficits caused by lesions of the cerebellum or TMS disruption of cerebellar function, and studying the potential functional meaning of simple- and complex-spike discharges in Purkinje cells during the elaboration of skillful voluntary movements. In most cases, control mechanisms have been studied in both monkeys and humans.
Load compensation task.
In Gilbert and Thach’s (
1977
) test, a monkey was trained to hold a lever stationary in the face of flexion and extension wrist forces developed by a torque motor. A change of load at some unpredictable time induced complex spikes in Purkinje cells located in the intermediate cortex of lobules III to V. These spikes occurred just after the load switch (at 50–150 milliseconds), and they apparently represented errors caused by the sudden change. Simple-spike discharge in these Purkinje cells changed inversely to their complex-spike discharge, and it remained decreased after the latter discharge had returned to normal. These observations provided early experimental evidence supporting Albus’ (
1971
) theoretical hypothesis (
Chapters 3
,
9
).
Reaction time.
“Simple reaction time” is the time taken from the presentation of a sensory stimulus (typically light or sound) to a subsequent behavioral response (typically pressing a button). It is considered to be an index of the speed of information processing in the relevant sensory-motor pathways. This time in college-age individuals is ~160 milliseconds for an auditory stimulus and ~190 milliseconds for a visual stimulus (
Brebner and Welford, 1980
). In a “serial reaction time” task, now widely used to measure the acquired degree of skill, a visual cue can appear at any one of four positions (1–4) arranged horizontally on a computer screen. When a cue appears as the go signal, the testee taps the appropriate button on a response pad, thereby ending the trial. The trials are repeated in 5 blocks of 100 trials/block. In blocks 1 and 5, the go signals are presented in random order. In blocks 2 through 4, the go signals represent a sequence of ten cues whose order (e.g., 2-3-1-4-3-2-4-1-3-4-2-1) is repeated ten times in each block of trials. The duration of each trial, defined by the participant’s response time, is the primary task measure, and its gradual shortening during the task is considered to be an index of learning (see, e.g.,
Gómez-Beldarrain et al., 1998
;
Robertson, 2007
).
In a group of normal subjects performing a serial reaction time task, repeated TMS (rTMS) over the lateral cerebellum significantly impaired learning. This interference with right cerebellar hemisphere activity induced a significant decrease in learning regardless of the hand used to perform the task, whereas left cerebellar hemisphere activity seemed to be more linked to learning using the ipsilateral hand (
Torriero et al., 2004
). In contrast, patients with single, unilateral vascular lesions in the territory of the posterior-inferior or superior cerebellar artery were examined as compared to age- and sex-matched control subjects in a one-handed version of a serial reaction time task. Learning was impaired for the hand ipsilateral to the lesion, but it was normal for the contralateral hand (
Gómez-Beldarrain et al., 1998
). The lateral cerebellar hemisphere thus appears to be involved in learning processes for a serial reaction time task for only the ipsilateral hand. However, another study on patients with unilateral focal cerebellar lesions reported that these patients were defective in learning the task with both hands (
Molinari et al., 1997
). Clearly, the interesting problem of functional lateralization in the cerebellar hemispheres is still an open issue.
The well-known finger-to-nose test of cerebellar dysmetria was reproduced when a monkey wearing wedge prisms was trained to try to reach a target by hand. Adaptation in the reaching depended critically on visual information about errors consequent to the movement at its end point. The effect of this visual information on consequence errors was largest at 0–10 milliseconds, and it decayed significantly in 50–100 milliseconds (
Kitazawa and Yin, 2002
). Thus, this paradigm represents typical consequence error-based learning. In another study by Kitazawa and his colleagues, a monkey was trained to perform short-lasting reaching (~200 milliseconds in duration) to touch a visual target, which appeared at random locations on a screen. The monkey saw its hand and fingers and the target before and after completion of the movement, but the movement itself was performed without visual feedback. In this case, Purkinje cells in cerebellar lobules HIV–HVI exhibited multiply timed climbing fiber responses at three different stages of the movement (first, second, and third responses). The third response occurred at the end point of the movement, apparently representing visually perceived deviations between the target and the reaching finger’s end position. However, the first and second Purkinje cell responses appeared too early to be interpreted similarly (
Kitazawa et al., 1998
). They apparently arose within the neuronal circuit controlling the hand-reaching movement (see
Chapter 15
, “
Internal Models for Voluntary Motor Control
,” for further details).
Reaching experiments have also been performed in cats to test the unique function of a set of propriospinal interneurons that have been studied extensively by Anders Lundberg and his colleagues (
Chapter 1
, “
Neuronal Circuitry: The Key to Unlocking the Brain
”). These interneurons, which are located in the C3–C4 spinal segments, receive major excitatory inputs from the corticospinal tract and also from the rubro-, reticulo-, and tectospinal tracts. The cells mediate their converged input, and that from other sources, to forelimb motoneurons (
Illert et al., 1977
,
1978
). Importantly, these C3–C4 propriospinal neurons have ascending collaterals to the lateral reticular nucleus (
Chapter 6
, “
Pre- and Post-Cerebellar Cortex Neurons
”), a major mossy fiber source for the cerebellum (
Illert and Lundberg, 1978
). The neurons thus appear to control forelimb movements while receiving adaptive modulation from the cerebellum. To test this hypothesis, cats were trained to make a fast, visually guided reaching movement toward a small tube containing a morsel of food, while the trajectory and velocity of the wrist movement were being monitored. A set of two or three such tubes were positioned in front of the animal at shoulder level, and the correct tube for retrieving the food was indicated by light-emitting diodes. The light was randomly switched off in one tube after a variable delay and switched on in one of the other tubes. The cat initiated a fast correction of the movement trajectory when switching from the left to the right tube. The switching latencies observed were as short as 40–60 milliseconds (
Pettersson et al., 1997
). After delays for retinal processing and electromechanical
delays in the muscles were subtracted, it seemed likely that within a time span of less than 20 milliseconds, information was processed to estimate the amount of correction needed to reach the new target position and give new command signals via subcortical systems to the propriospinal neurons (
Alstermark et al., 2007
). How the cerebellar circuit computes such new signals is still an open question (see
Chapter 18
, “
Concluding Thoughts
,”
Section 3
).
In another study, Purkinje cell activity was observed in cats trained to use the left paw to track a visual target moving from the left to the right in front of the animal. For successful reaching, the cat had to predict where the target would be when it received the go signal (brightening of a light-emitting diode) for initiation of the tracking movement. Some (23/50) of the Purkinje cells sampled in the lateral cerebellar D
1
-zone of crus I displayed an increase or a decrease in simple-spike discharge rate in response to the onset of target movement. Other of the cells (14/33 tested) responded to the go signal with modulation, the great majority (12/14) with an increase in discharge rate. The results were interpreted to suggest that these Purkinje cells were predicting the initial movement of the target (
Miles et al., 2006
).