The Cerebellum: Brain for an Implicit Self (3 page)
Read The Cerebellum: Brain for an Implicit Self Online
Authors: Masao Ito
Tags: #Science, #Life Sciences, #Medical, #Biology, #Neurology, #Neuroscience

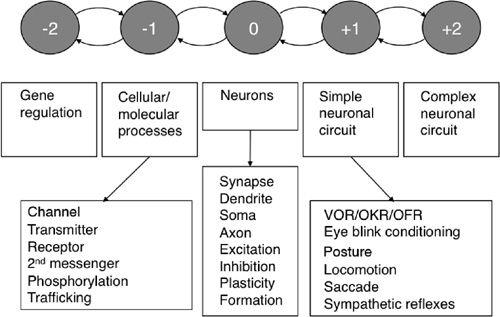
These processes can be considered as a long chain of decomposition-reconstruction events. By successively linking hierarchical levels, neuroscience research can trace the long pathway of evolution, from organic molecules to the cells of multicellular organisms, and eventually to the differentiated and diversified neurons that constitute simple neuronal circuits. In addition, evolutionary processes starting from simple neuronal circuits gradually led to the development of increasingly complex circuits and finally the human CNS. The fields of many subdisciplines of neuroscience are found at specific levels of the hierarchy. For one to understand the mechanisms and roles of neuronal circuits in the CNS, consistent and sustained effort is required to link coherently all levels of the hierarchy centering
around neuronal circuits that extend to cells and molecules on one hand, and to complex networks and systems on the other. In later chapters, we will see how far the cerebellum has been decomposed and reconstructed using these general methodologies.
The concept of the “neuron” was established over a century ago as the unitary component of neuronal circuits. Ramón y Cajal (1852–1934), hereafter shortened to “Cajal,” presented clear evidence for this in 1888, when referring to the relationship between Purkinje and basket cells in the cerebellum (see below and Color Plate IV) (Lopez-Munoz et al., 2006). Heinrich von Waldeyer-Hartz (1836–1921) formally proposed the neuron theory in 1891. Also, near the end of the nineteenth century, Sherrington and Michael Foster (1836–1907) coined the term “synapse” and spotlighted it as a key structure of the CNS. Since then, neurons and synapses have been the major targets of neuroscience investigations. All neurons commonly have somata extruding axons and dendrites (except for dorsal root ganglion cells, which have no dendrites). Dendrites not only expand the membrane area to accommodate many hundreds of synapses, but they also have finely
compartmentalized functions (
Hausser and Mel, 2003
). On the other hand, different types of neurons are distinguished by their characteristic morphology, spike activities, synaptic actions (excitatory or inhibitory), and synaptic receptiveness (chemical or electrical). Subcellular structures such as postsynaptic density (PSD), cytoskeleton, endoplasmic reticulum, Golgi organ, and mitochondrion support these neuronal functions. Signal transduction involves various transmitters, modulators, receptors (ionotropic or metabotropic, or both), and second messengers. For these molecular mechanisms of neurons, numerous proteins, glycoproteins, and lipids, and their genes play essential roles.
Numerous neurons in the CNS assemble to form a structure called a “nucleus.” In certain areas of the brain and spinal cord (e.g., the superior colliculus, cerebellar cortex, hippocampal cortex, cerebral neocortex), different types of neurons regularly assemble to form a multilayered network. Donald Hebb (1904–1985) proposed the concept of “neuron assembly,” that is, a collation of neurons interconnected by synapses, in which the connectivity is modifiable according to experienced activities (
Hebb, 1949
). A famous proposal by Hebb is that the connection between two neurons firing synchronously is strengthened. Because of this “Hebbian” type of synaptic plasticity; a neuronal assembly can change its circuitry structure (corresponding to memory) and consequently modify its input-output relationships (corresponding to learning), as dependent on experienced activities.
In an effort to verify Hebb’s concept of neuron assembly, Frank Rosenblatt (1928–1971) constructed a model network named a “simple perceptron.” It consisted of three layers of neurons connected in one direction, from the sensory cell layer to the association cell layer, to the response cell layer (
Figure 6
). Connections from the first to the second layer were fixed, whereas those from the second to the third layer were modifiable according to the instruction of an outside “teacher.” The teacher increased the weight of connection at all junctions transmitting signals from the second to the third layer when the simple perceptron responded correctly to sensory stimuli. The teacher decreased the weight at all second-to-third layer connections transmitting signals when the response was incorrect. When this training process was repeated, the simple perceptron improved its performance toward a success rate of 100%. This was the first man-made machine capable of learning. Ten years later, a counterpart of the simple perceptron was found in the cerebellum (see
Chapters 3
and
9
). The simple perceptron exemplified the success of the constructive approach (i.e., to understand by construction) for clarifying the operation of neuronal networks in the CNS.
Figure 6. The simple perceptron model of the cerebellum.
This figure is self-explanatory. See the text for further details on the operation of a simple perceptron. Abbreviation: CF, climbing fiber.

Twenty-four years after the construction of the simple perceptron, another form of multilayered neuronal assembly was proposed. It is usually called a “neurocomputer” (
Rumelhart et al., 1986
), in which errors were estimated by comparing the output of the third layer with a preset goal and were back propagated to the third-layer neurons. The errors acted on the junctions on the third-layer neurons formed with second-layer (hidden layer) neurons, and modified the efficacy of transmission from second-layer to third-layer neurons. The neurocomputer is often applied to model information processing in hippocampal and neocortical networks.
Local networks are interconnected globally throughout the CNS to form neural “systems.” A major type of such a system has the general form of a “control system,” which consists of a “controller (g)” acting on a “controlled object (G)” (
Figure 7A
). The controller receives input instruction that provides information about the nature of the required output (e.g., the goal, the trajectory of a movement). In turn, the controller generates command signals that drive the controlled object to respond appropriately. The controller may receive information about the performance of the controlled object (
Figure 7A
, feedback control), or it may operate without peripheral information (
Figure 7B
, feedforward control). The goal
of a control system is to generate output responses identical to the input instruction. This can be achieved in a feedback control system if g is sufficiently larger than G, but in a feedforward control system, g needs to equal 1/G (
Figure 7B
). As emphasized by Baev (
1999
), this basic control system concept applies to various levels of organization within the CNS: in this monograph from reflexes to isolated voluntary movements and finally to coordinated motor actions. In addition, the concept is applied formalistically to cognitive functions.
Figure 7. The fundamental structures of a control system.
(A) A basic feedback control system. (B) A basic feedforward control system. (C) An adaptive control system equipped with an adaptive mechanism. This schematic applies to the cerebellar control of reflexes.

In recent years, modern control theory studies have opened the new fields of “adaptive control” and “model-based control.” In adaptive control, the controller is equipped with an adaptive mechanism to constitute an adaptive controller, which learns how to perform effectively in a given situation by altering its performance to match ever-changing environments. When a mechanism is attached to a feedforward controller, their overall input-output relationship f should be adjusted to 1/G (
Figure 7C
). On the other hand, model-based control was developed for robotic
arm control (
An et al., 1988
), and it has opened a new field of computational neuroscience for research on the cerebellum (
Kawato et al., 1987
).
In the model-based control, a feedforward control system (
Figure 7B
) is attached with one of the two types of internal models, “forward” and “inverse” (
Figure 8A
,
B
). An internal forward model simulates the kinematics of a controlled object, whereas an internal inverse model simulates the dynamics or kinetics of them (for a definition, see
Chapter 15
, “
Internal Models for Voluntary Motor Control
”). Internal forward models support the controller by predicting the state of the system during actual actions. On the other hand, internal inverse models map the relationship between intended actions (or goals) and the motor command to bring about the action. An internal inverse model uses the desired position of the body as inputs to estimate the necessary motor commands, which would transform the current position into the desired one. An adaptive mechanism is involved to secure close simulation by these models. Such models may be formed in various parts of the CNS including, in particular, the elaborate neuronal networks of the cerebellar and cerebral cortices. Hereafter, models formed in the cerebellum and cerebral cortex will be called “cerebellar internal models” and “cerebral cortical models,” respectively.
Figure 8. General forms of model-based control systems.
(A) Internal forward model (G’) simulates the input-output relationship of the controlled object (G) and is inserted between the output and input of the controller (g). (B) Internal inverse model (1/G) simulates the output-input relationship of the controlled object (G) and is inserted between the input instruction and output response of the controller (g).

The most fundamental control system structure in the CNS is an individual reflex operating via the spinal cord or brainstem. A reflex is performed by sequential activities in a neuronal circuit that connects serially sensory receptor cells, an afferent path, a reflex center, an efferent path, and an effector (a muscle, set of muscles, or a secretory gland or glands). This is typically exemplified by the stretch reflex, in which motoneurons maintain the length of a muscle constant by using feedback from muscle spindles (
Chapter 11
, “
Somatic and Autonomic Reflexes
”). In this case, a group of motoneurons and associated segmental interneurons constitute a controller, whereas the motor apparatus composed of the muscle(s) and a joint provides the controlled object. Numerous reflexes of various types operate in the spinal cord and brainstem to control simple somatic and visceral functions of a living body. The operation of reflexes is usually automatic—that is, it does not reach the level of conscious awareness—but in ever-changing environments it is indeed modifiable by use of adaptive mechanisms of the cerebellum (
Chapters 10
–
12
).