Connectome (27 page)
Authors: Sebastian Seung

Â
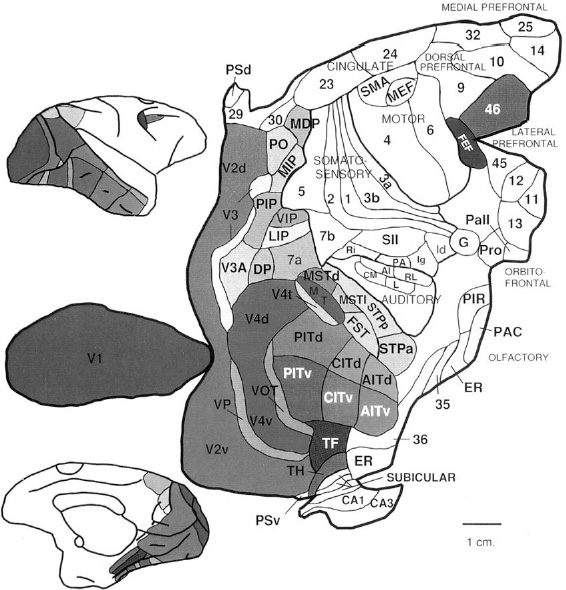
A modern map of a monkey brain (see Figure 39) brings back pleasant memories of my father's atlas. Its colorings bear mysterious acronyms, and sharp corners punctuate its gentle curves. But maps are not always so charming. Let's not forget that armies have clashed over lines drawn on them. Likewise, neuroanatomists have waged bitter intellectual battles over the boundaries of brain regions.
Â
 Â
Â
Â
Figure 39. Map of the rhesus monkey cortex laid out flat
Â
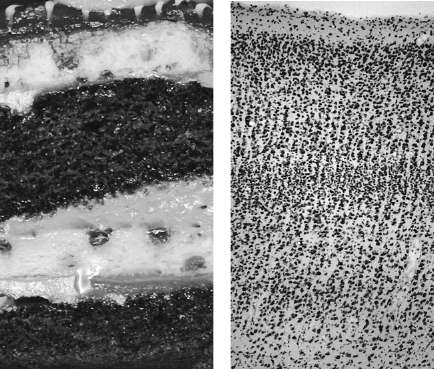
We already encountered Korbinian Brodmann's map of the cortex. How exactly did he create it? The Golgi stain allowed neuroanatomists to see the branches of neurons clearly. Brodmann used another important stain, invented by the German neuroanatomist Franz Nissl, which spared the branches but made all cell bodies visible in a microscope. The stain reveals that the cortex (Figure 40, right) resembles a layer cake (left). Cell bodies are arranged in parallel layers that run throughout the entire cortical sheet. (The white spaces between cell bodies are filled with entangled neurites, which are not marked by the Nissl stain.) The boundaries in the cortex are not as distinct as those of the cake, but expert neuroanatomists can make out six layers
. This piece of cortical cake, less than one millimeter wide, was cut from a particular location in the cortical sheet. In general, pieces from different locations have different layerings. Brodmann peered into his microscope to see these differences, and used them to divide the cortex into forty-three areas. He claimed that the layering was uniform
at every location within one of his areas, changing only at the boundaries between areas.
Â
 Â
Â
Â
Figure 40. Layers: cake
(left)
and Brodmann area 17, also known as V1 or primary visual cortex
(right)
Â
Brodmann's map of the cortex may be famous, but it shouldn't be taken as gospel truth. There have been plenty of other contenders. Brodmann's colleagues in Berlin, the husband-and-wife team of Oskar and Cécile Vogt,
used a different kind of stain to divide the cortex into two hundred areas. Still other maps were proposed by Alfred Campbell working in Liverpool, Sir Grafton Smith
in Cairo, and Constantin von Economo and Georg Koskinas in Vienna. Some borders were recognized by all researchers, but others sparked discord. In a 1951 book Percival Bailey
and Gerhardt von Bonin erased most of the borders of their predecessors, leaving just a handful of large regions.
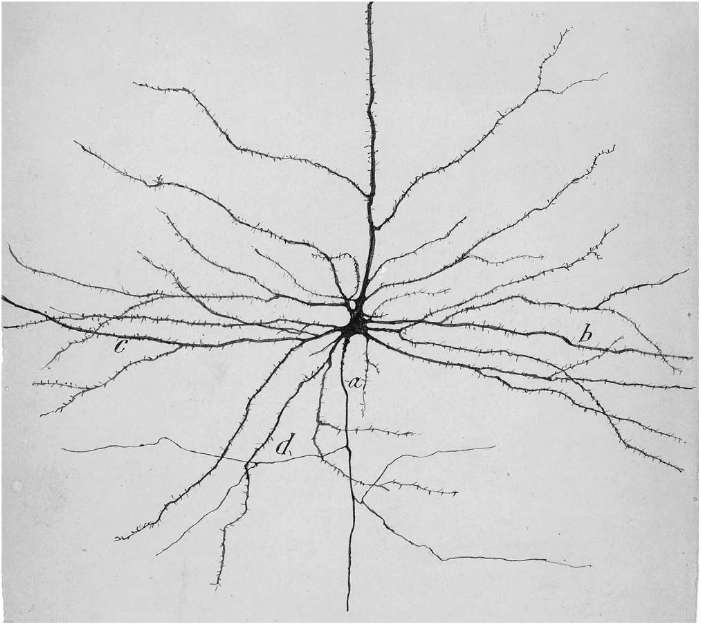
Even worse controversies have plagued Cajal's program to classify neurons into types. He did this based on their appearance, much as a nineteenth-century naturalist would have classified different species of butterflies. One of his favorite neurons was the pyramidal cell. He called it the “psychic cell,” not because he believed in the occult, but because he thought that this type of neuron played an important role in the highest functions of the psyche. In Figure 41, a drawing by Cajal himself, you can see the defining features of this neuron type: the roughly pyramidal shape of its cell body, the thornlike spines protruding from its dendrites, and the long axon traveling far away from the cell body. (The axon is directed downward in this image, descending into the brain. The most prominent “apical” dendrite leaves the apex of the pyramid and travels upward, toward the surface of the cortex.)
Â
 Â
Â
Â
Figure 41. Drawing of a pyramidal neuron by Cajal
Â
The pyramidal cell is the most common type of neuron in the cortex. Cajal observed other cortical neurons that had shorter axons, and smooth rather than spiny dendrites. The shapes of nonpyramidal neurons were more diverse, so they were divided into more types, which earned picturesque names such as “double bouquet cell.”
Cajal classified neurons into types all over the brain, not just in the cortex. This alternative method of carving up the brain was much more complex than Brodmann's, because every brain region contains many neuron types. Furthermore, the types in each region are intermingled, like different ethnic groups living in the same country. Cajal could not complete the task in his lifetime, and even today this enterprise has only just begun. We still don't know how many types there are, though we know the number is large. The brain is more like a tropical rainforest, which contains hundreds of species, than a coniferous forest with perhaps a single species of pine tree. One expert has estimated that there are hundreds of neuron types
in the cortex alone. Neuroscientists continue to argue
over their classification.
Their disagreements are a sign of a more fundamental problem: It's not even clear how to properly define the concepts of “brain region” and “neuron type.” In Plato's dialogue
Phaedrus,
Socrates recommends “division . . . according to the natural formation, where the joint is, not breaking any part as a bad carver might.” This metaphor vividly compares the intellectual challenge of taxonomy to the more visceral activity of cutting poultry into pieces. Anatomists follow Socrates literally, dividing the body by naming its bones, muscles, organs, and so on. Does his advice also make sense for the brain?
“Carving nature at its joints” means cutting in places where connections are weakest. It doesn't take a professional to divide the brain into two hemispheres by cutting the corpus callosum. But most brain regions are not so obvious. The boundaries between cortical areas don't seem like “joints” in the cortical sheet. A great many wires
extend across them, connecting neurons on either side.
Of course, we've already carved the brain into extremely fine pieces: individual neurons. No one argues over whether these divisions are objectively defined, now that the debate between Golgi and Cajal has been settled. But as I mentioned in relation to research on Parkinson's disease, it's also useful to divide the brain more coarsely, into regions and neuron types. How can we make these divisions more accurate?
I believe that connectomes will give us new and better ways to divide the brain. We will have to follow Socrates less literally. Unlike poultry,
connectomes will be carved in a more abstract way, by classifying neurons based on their connectivity. This approach was previously used to divide the three hundred neurons of
C. elegans
into over one hundred types.
The researchers followed a basic principle: If two neurons are connected to similar or analogous partners, they should be grouped in the same type. Some types are trivial, consisting of only a neuron and its twin on the opposite side of the body. The left and right neurons of such a pair are connected to analogous neurons, much as your left arm is connected to your left shoulder and your right arm to your right shoulder. Other types are less trivial, containing up to thirteen neurons with similar connectivity.
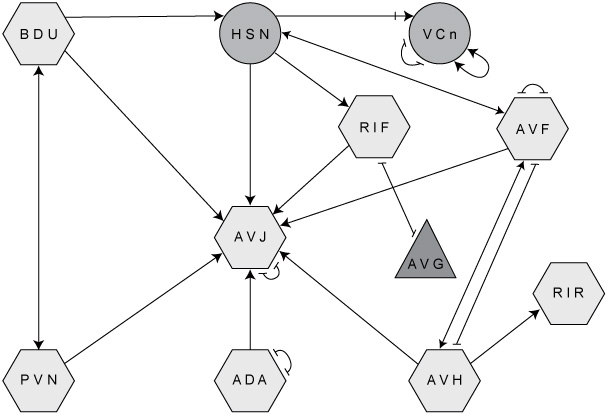
Using neuron types, we can simplify the diagram of the
C. elegans
connectome shown in the Introduction (Figure 3). Collapse all neurons of one type
into a single node, and repeat this for all types. Figure 42 shows a portion of the result. Each three-letter acronym specifies a neuron type involved in egg-laying behavior. “VCn,” for example, stands for the neurons VC1 through VC6, which control the vulval muscles. The lines between the nodes represent connections between neuron types rather than neurons, so we might call the diagram a
neuron type connectome.
Â
 Â
Â
Â
Figure 42. Portion of the “reduced” connectome (with neurons grouped by type) of C. elegans
Â
This example shows that carving a connectome not only yields neuron types but tells us how they are connected. Neuroscientists would like to do the same for the retina. The connections between the five broad neuron classes are already known. For example, horizontal cells receive excitatory synapses from photoreceptors, and send back inhibitory synapses. They also make electrical synapses with each other. But recall that the five classes are divided into more than fifty neuron types. Their connectivity is mostly unknown but could be discovered by finding and carving the neuronal connectome of the retina.
It's worth noting that this approach is different from the classical one. Cajal first defined neuron types by shape and location, and then moved on to investigate their connections. Here I'm proposing to turn this around, starting from connectivity and working backward to define neuron types.
This approach, though different, can still be viewed as a refinement of Cajal's if we regard shape and location as proxies for connectivity. To understand why, imagine looking at two neurons. Each neuron extends its branches over some region. If the two regions are completely separate from each other, for genetic or other reasons, then there is no way for the neurons to be connected. Contact is a prerequisite for connection, and contact is governed by location and shape.
If shape and location are so closely related to connectivity, why is connectivity the better approach? The answer is the connectionist mantra, “The function of a neuron is chiefly defined by its connections with other neurons.” Connections are directly related
to function, while shape and location are only indirectly so.
A similar strategy can be applied to divide the brain more coarsely into regions rather than neuron types. In the discussion of rewiring I mentioned that each cortical area possesses a unique “connectional fingerprint,” or pattern of connection with other cortical areas as well as regions outside the cortex. We could turn this around and use it to define cortical areas. If we carve a connectome into groups of contiguous neurons, so that each group shares a common connectional fingerprint, we should end up with brain regions. (We'd have to constrain the groups to not overlap in space, or else they might end up being intermingled neuron types rather than spatially distinct regions.)
How is this related to Brodmann's use of layering to define cortical areas? Again, layering should be regarded as a proxy for connectivity. For example, areas 17 and 18 differ by the thickness of layer 4, because of their different connectivity. Layer 4 of area 17
is swollen with many neurons that receive connections from pathways originating in the eyes. The adjacent area 18 doesn't receive such axons, so its layer 4 is not as large.