Connectome (28 page)
Authors: Sebastian Seung

If layering is so closely related to connectivity, why should we prefer a definition based on the latter? Once again, it's because the layering is less fundamental. The fact that visual pathways lead to area 17 tells us immediately that its function is visual. The fact that area 17 has a thicker layer 4
is only indirectly related to its function.
Brodmann relied on cortical layering and Cajal on neural shape and location. Although more sophisticated than size, these properties were still only crude substitutes for what really matters: connectivity. More than a century later, we should be able to dispense with the proxies and work directly with connectomes.
Â
I've argued that the ideal way of dividing a brain is to carve its connectome. As a bonus, we also learn how the divisions are connected with each other, obtaining a regional or neuron type connectome.
How can these simplified versions of the neuronal connectome help us understand the brain?
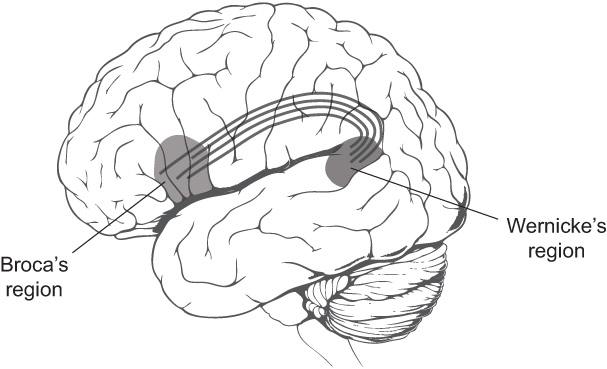
The importance of regional connections was recognized as early as the nineteenth century, when Wernicke hypothesized a bundle of long axons connecting Broca's and Wernicke's regions. Damage to the bundle, while leaving both speech comprehension and production intact, would make it impossible to repeat words after hearing someone else speak them. Wernicke's region could receive the words but could not relay them to Broca's region to be spoken aloud. Because this hypothetical disorder was due to loss of signal conduction, Wernicke called it
conduction aphasia.
Patients with these symptoms were later discovered, confirming Wernicke's prediction. Furthermore, neuroanatomists identified the hypothetical connection between Broca's and Wernicke's regions as a bundle of axons called the arcuate fasciculus (see Figure 43).
Â
 Â
Â
Â
Figure 43. A bundle of axons connecting Broca's and Wernicke's regions
Â
The BrocaâWernicke model of language shows how one might go about using the regional connectome once it's found. Associate every brain region with an elementary mental function, such as speech comprehension or production. Then explain more complex mental functions, like speech repetition, as combinations of elementary functions. These are carried out by cooperation between multiple regions, which is mediated by regional connections.
Neurologists use this conceptual framework to diagnose patients with brain damage. Damage to a region impairs the corresponding elementary function. Damage to a
connection
impairs complex functions requiring cooperation between regions. Because this paradigm includes connections and allows for distributed functions, it goes beyond localizationism. It is sometimes called connectionism, though it's a different flavor from the neural
connectionism introduced earlier. We can also imagine a connectionism based on neuron types. This kind of brain model would be more sophisticated than the neurologists', and far more challenging to construct, because neuron types and their connections are so numerous.
But in the immediate future, a regional connectome seems like the most useful kind for psychologists and neurologists. Olaf Sporns and his colleagues
pointed this out when they originally coined the term
connectome
in a 2005 paper. You may have heard of the $30 million Human Connectome Project, which was launched in 2010 by the U.S. National Institutes of Health (NIH). Most people don't realize that this project is only about regional connectomes, and has nothing to do with neuronal connectomes.
I personally spend more time focused on neurons than on regions, but I agree with Sporns and his colleagues about the importance of finding regional connectomes. My only disagreement has to do with the methods. In my view, we need to see neurons to find regional connectomes. In other words, I am still a neuronal chauvinistâbut about means rather than ends.
I believe it would be best to find regional connectomes by carving neuronal connectomes, though I admit that this strategy is idealistic at present. In the near term it will be practical only for very small brains, not for human brains. That's why the Human Connectome Project is attempting a shortcut: finding regional connectomes using MRI. As I will explain later, this imaging method is bound to encounter difficulties, because of its limited spatial resolution. In Chapter 12 I will propose an alternative shortcut to finding regional connectomes, one that might be practical in the near future without cutting as many corners. A similar shortcut could also help us find neuron type connectomes.
Â
Not every neuroscientist is convinced that we should put more effort into dividing the brain; some believe that our maps are already good enough. To counter this idea, let's take a closer look at the BrocaâWernicke model of language. It sounds so successful in the textbooks, but the true story is messier.
In Broca's original patient, the brain lesion covered much more than Broca's region, extending into surrounding cortical areas as well as regions below the cortex. It turns out that a lesion of Broca's region alone does not produce Broca's aphasia, and lesions that spare
Broca's region can produce Broca's aphasia. The regional basis of Wernicke's aphasia is similarly murky. Furthermore, the double dissociation of speech production and comprehension is not as clean as the textbooks make it sound. For example, Broca's aphasia is usually accompanied by problems with comprehension of sentences. Consistent with these clinical findings, recent fMRI studies have shown that language is less localized than previously
thought, involving cortical and subcortical areas beyond those of Broca and Wernicke. Clinical studies do not support the traditional story that conduction aphasia is caused by lesions of the arcuate fasciculus. More embarrassingly, some researchers now deny that the arcuate fasciculus
connects Broca's and Wernicke's regions, even though we've believed it does for over a century. Some neuroscientists have found other pathways that do
connect these regions.
For all these reasons, language researchers are struggling to formulate a replacement for the BrocaâWernicke model.
The new model will have to include additional cortical areas, as well as brain regions outside the cortex, and will have to explain a more complex array of linguistic abilities than the simplistic duo of speech comprehension and production. Everyone agrees on the need for an improved model, but there is no consensus on how to find it. I don't presume to know, but I am sure that better maps of the brain would be helpful.
Dividing the brain has historically been more art than science. Like a physician's divining a disease from a constellation of symptoms or a judge's compromise between multiple legal precedents, dividing the brain has never been reduced to a simple formula. Some of the boundaries between brain regions are no doubt arbitrary, the result of historical accidents and errors of neuroanatomists. Like globes and atlases, our maps of the brain do not represent objective, timeless truth. Sometimes new regions are created, or the borders between them may shift. Disputes over borders can erupt in acrimonious debate between scientists, ideally settled peacefully by the patient negotiations of committees.
We should not become complacent with this state of affairs. Our current maps of the brain may not be as poor as world maps from centuries ago, which look almost laughable to our modern eyes, but there is plenty of room for improvement. Maps by themselves will not tell us how brain regions contribute to mental function. They will, however, accelerate research by providing a firm foundation on which to stand.
My emphasis on structural criteria for dividing the brain will seem strange to a contemporary neuroscientist, who is used to combining them with functional criteria. But this kind of emphasis is commonplace in the rest of biology. The organs of the body were known as structural units long before their roles were understood, and can be identified by a naive observer with no knowledge of function. Similarly, the organelles of a cell were observed in the microscope long before it was known that the nucleus contains the genetic information and that the Golgi apparatus packages proteins and other biomolecules before sending them to their proper destination.
In general, biological units are both structural and functional entities, but they are typically identified first by structure; their functions are figured out later. Brain regions and neuron types should be the same way. Neuroscientists following Brodmann and Cajal have long pursued the structural approach to dividing the brain but have achieved only partial success. The problem is not that the approach is fundamentally flawed; rather, it is that our techniques for measuring brain structure have been inadequate. Any division of the brain is only as good as the data on which it is based. By providing dramatically better data about structure, connectomics will allow a more objective division of the brain, and by extension the mind.
Identifying cortical areas by examining the symptoms of brain damage is analogous to the way that the Austrian monk Gregor Mendel identified genes in the 1860s. His experiments on the interbreeding of plants showed that the inheritance of certain traits (now called Mendelian) was controlled by variations in a single unit, later called a gene. In his simple picture, traits and genes had a one-to-one correspondence. But now we know that most traits are not Mendelian. Most traits can be affected by many genes, and one gene may affect many traits. This is because a gene encodes for a protein, which can perform many tasks.
Similarly, localizationism attempted to establish a one-to-one correspondence between mental functions and cortical areas. But it turns out that most mental functions require the cooperation of multiple cortical areas, and most cortical areas participate in multiple mental functions. This makes it problematic to use functional criteria to define cortical areas. The right strategy is to identify the areas by structural criteria and then understand how the interactions between areas give rise to mental functions. This approach will become practical as our technologies improve.
We expect to find the same regions and neuron types in all normal brains. The regional and neuron type connectomes are likely to vary little across normal individuals, and are likely to be highly determined by genes. As I mentioned earlier, genes guide the growth of the branches of neurons, thereby influencing the neuron type connectome. Scientists are also identifying genes that control the formation of cortical areas.
Your mind and mine may be similar because our brain regions and neuron types are connected in the same ways.
In contrast, neuronal connectomes will vary greatly across individuals and will be strongly influenced by experiences. These are the connectomes we must study if we want to understand human uniqueness. And we should examine them for traces of the past, for what could be more integral to our uniqueness than our own memories?
I Likened the task of seeing connectomes to navigating the twisting passages of the Labyrinth. According to legend, this structure was located near the palace of King Minos at Knossos on the island of Crete. In 1900 Knossos yielded a second metaphor for the brain. Hundreds of clay tablets were excavated from the ancient ruins. Their discoverer, the British archaeologist Arthur Evans, could not read them, for they were inscribed in an unknown language. For decades the tablets remained unintelligible, and their mysterious script became known as Linear B. Finally, in the 1950s, Michael Ventris and John Chadwick
succeeded in decoding Linear B, and the tablets' meaning was revealed.
Once we can see connectomes and carve them up into pieces, the next challenge will be to decode them. Will we learn to understand their language? Or will their patterns of connectivity merely tantalize, refusing to give up their secrets? Decoding Linear B took half a century, but at least Ventris and Chadwick succeeded in the end. Similar attempts have failed for a number of lost languages.
Linear A, the script used in ancient Crete before Linear B, remains incomprehensible. The Indus script of ancient Pakistan, the Zapotec writing system of ancient Mexico, and the Rongorongo glyphs of Easter Island have also eluded decipherment.
What exactly does it mean to decode connectomes? Sometimes it's easier to understand a concept if you consider its most extreme version. For a thought experiment, let's imagine that you are living in the far future. Medicine has become very advanced, but, alas, your great-great-grandmother finally dies (at age 213). You take her to a facility that slices up her brain, images the slices, finds her connectome, and hands you a little electronic stick containing the data. After you get home, you are feeling sad because you miss talking to her. (She was your favorite great-great-grandmother.) You place the stick in your computer
and ask it to recall some of her memories, and soon you are feeling better.
Will it ever be possible to read memories from connectomes? I proposed a similar thought experiment earlier, asking you to consider whether someone could read your perceptions and thoughts by measuring and decoding the spiking of every neuron in your brain. Some neuroscientists believe we could do this if our technologies for measuring spiking were advanced enough. Why do they think so? From the spiking of a Jennifer Aniston neuron we can already guess whether a person is perceiving Jen. Neuroscientists extrapolate from this small success that the spiking of all neurons would give us a complete picture of our thoughts and perceptions.