Connectome (32 page)
Authors: Sebastian Seung

Â
Figure 49. Collateral and main branches of a pyramidal neuron's axon
Â
As the axon dives down, it sends out side branches, called “collaterals,” which are for making synapses onto nearby neurons. But the main branch of the axon finally leaves the gray matter and enters the white matter to start its journey to other regions. In each of its destination regions, it forks out many branches to make connections with neurons there.
Some axons don't travel very far, reentering the gray matter close to where they started. But most axons of pyramidal neurons project to other regions in the cortex, some going as far as the other side of the brain. Some white-matter axonsâa small minorityâconnect the cortex with other structures in the brain, such as the cerebellum, the brainstem, or even the spinal cord. These axons make up less than one-tenth of the white matter. The cortex is highly self-centered, primarily “talking” with itself rather than the outside world.
Here's another way to think about it: If the axons and dendrites in the gray matter are like local streets, the axons of the white matter are like the superhighways of the brain. They are relatively wide and unbranched, and also extremely long. In fact, the total length of these axons is roughly 150,000 kilometers,
over a quarter of the distance from the Earth to the Moon. And herein lies the challenge: Finding the regional connectome requires tracing the journey of every axon in the white matter.
It seems like an impossible task, but it could be done by slicing and imaging all of the white matter and using computers to follow the path traveled by each axon in the images. The start and end points of every path would define a connection between two locations in the cortex. Is this approach too difficult to be practical? After all, the cerebral white matter is comparable in volume to the gray matter, and we are still struggling to reconstruct one cubic millimeter of that. Given this, it might seem outlandish to propose reconstructing hundreds of cubic centimeters of white matter. My proposal seems less crazy once you know that white-matter axons are visible at a lower resolution.
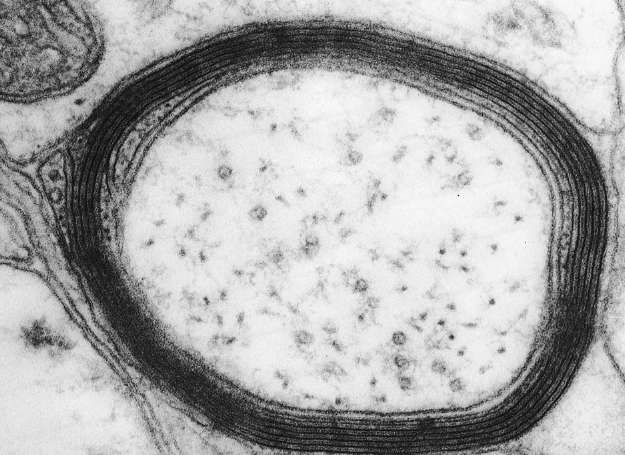
To understand why, take a look at the cross-sectional image shown in Figure 50. As axons exit the gray matter, most of them undergo an important transformationâthey become ensheathed by other cells that wrap around them repeatedly. Thus the brain not only wires itself up but also, amazingly, manages to wrap sheets of insulation around its “wires.” The sheets are made of a substance called myelin, which is composed mostly of fat molecules. It's those molecules that make white matter look white. (The epithet “fathead” may sound derogatory, but it's actually accurate for everyone.) Myelination speeds up
the propagation of spikes, which is important for transmitting signals quickly in large brains. Diseases of myelination, such as multiple sclerosis, have catastrophic effects on brain function.
Â
 Â
Â
Â
Figure 50. Cross-section of myelinated axon
Â
The myelinated axons of the white matter are much thicker (typically 1 micrometer) than the mostly unmyelinated axons of the gray matter. Furthermore, if we only care about finding regional connections, there's no need to see synapses. If an axon enters and branches
in a region of the gray matter, we can be almost certain that it makes synapses there, so tracing the “wires” of the white matter is enough for finding the regional connectome. If we restrict ourselves to myelinated axons, we could accomplish the job with serial light microscopy, which is similar to serial electron microscopy but employs thicker slices and produces images with lower resolution.
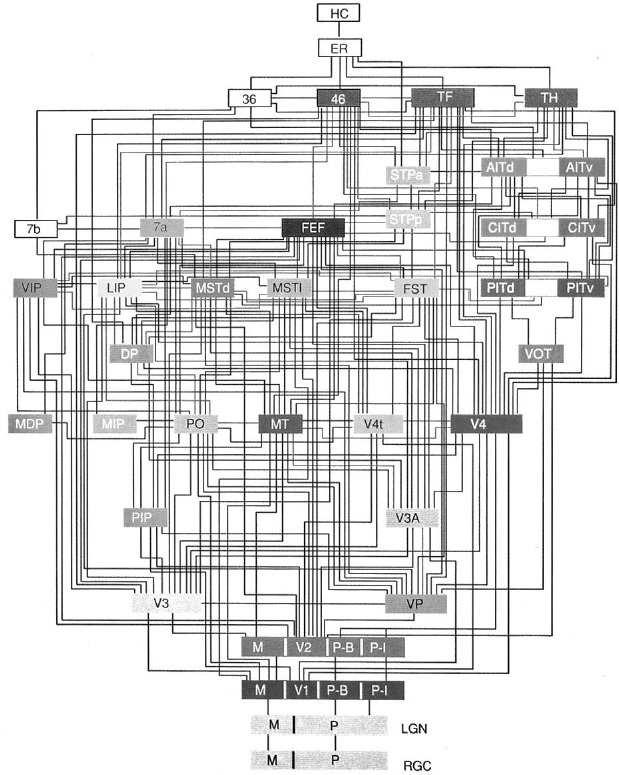
Of course, mapping white-matter axons is still a daunting technical challenge for a brain of human size. Studying white matter in smaller brains, such as those of rodents and nonhuman primates, is a good starting place. We can check the results by comparing them with those from older techniques for studying white-matter pathways in animals. These techniques were used to find connections between the visual areas of the monkey cortex, as shown in Figure 51. (The areas, but not their connections, were shown earlier.) Since the older techniques are not applicable to human brains, our own white matter has gone almost completely unexplored.
Â
 Â
Â
Â
Figure 51. Connections between visual areas of the rhesus monkey cortex (see Figure 39)
Â
The Human Connectome Project is already trying to find a map like the one in Figure 51 for the human brain using diffusion MRI (dMRI) rather than microscopy. Diffusion MRI is different from MRI, which is used to find the sizes of brain regions, or fMRI, which is used to measure their activations. Unfortunately, dMRI is subject to the same basic limitation as other forms of MRI: poor spatial resolution. MRI typically yields millimeter-scale resolution, which is not enough for seeing single neurons or axons. Given its poor resolution, how can dMRI hope to trace the wires in the white matter?
It turns out that white matter has an interesting feature that makes its structure simpler than that of gray matter. Have you ever forgotten to stir the spaghetti after dropping it into boiling water? You discover your mistake a few minutes later, when you see that some of the strands have stuck to each other to form bundles. This culinary embarrassment resembles white matter; gray matter is more like a bowl of fully entangled spaghetti.
When axons bundle like unstirred spaghetti, they form a “fiber tract” or a “white-matter pathway.” The bundles are similar to nerves, except that they run within the brain. Why do axons bundle? Well, why do so many people follow the same dirt paths through lawns? First, they are shortcuts, more efficient than the paved walkways installed by landscape designers. Second, there is a “follow the leader” effectâonce a few trailblazers have worn down the grass a bit, everyone else follows them, trampling it down completely. Similarly, axons take efficient paths through the white matter, assuming that it evolved to achieve wiring economy. Since an efficient solution is often unique, we'd expect axons sharing the same origin and destination to take the same path. Also, it's known that the first axons to grow during brain development often blaze the trail, providing chemical cues for other axons to follow.
Fiber tracts may be thick, even though a single axon is microscopically thin. The largest is the famous corpus callosum, the huge collection of axons that travel between the left and right hemispheres. Neuroanatomists in the nineteenth century discovered other large tracts through naked-eye dissection of the brain. Diffusion MRI is an exciting
advance, because it's a way of tracing white-matter pathways in the living brain. It computes an arrow at every location that indicates the orientation of the axons there. By connecting these arrows, it's possible to trace the paths of axonal bundles. In one notable success, dMRI has uncovered white-matter pathways connecting Broca's and Wernicke's regions, other than the classical one in the arcuate fasciculus. As I mentioned earlier, such discoveries are sparking revisions
of the BrocaâWernicke model of language.
Such stories are encouraging, but dMRI also has limitations. Because of its poor spatial resolution, dMRI has difficulty following thin fiber tracts. And even thick tracts can be problematic if they intersect and their individual axons become intermingled. Think of this crossing as a chaotic traffic intersection packed with pedestrians, bicyclists, animals, and carsâyou have to watch carefully to see whether any particular traveler goes straight or turns. Similarly, once axons enter the region where two bundles intersect, it's difficult to see, using dMRI, where they end up. The only foolproof way of mapping the white matter is to use a method that can trace individual axons, like the one I've proposed here.
Mapping regional connectomes is already problematic with dMRI; the method is even more ill-suited for neuronal and neuron type connectomes. Of course, dMRI has the important advantage that it can be performed on a living brain. At the very least it will detect gross connectopathies, like a missing corpus callosum. Since dMRI can be used quickly and conveniently to study many living brains, it will find correlations between mental disorders and brain connectivity. But these correlations might remain weak, just like the earlier phrenological ones.
MRI experts are continuing to improve resolution, but the rate of improvement is not that fast, and there is a long way to go. Roughly speaking, the current resolution of dMRI is a thousand times worse than light microscopy, which in turn is a thousand times worse than electron microscopy. Inventors might create better noninvasive imaging methods than MRI. But let's not forget that seeing through the skull into the interior of a living brain is fundamentally more challenging than chopping up a dead brain and examining the pieces with a microscope. Microscopy already delivers the resolution we need to find connectomes; we just have to scale it up to handle larger volumes. In contrast, MRI requires breakthroughs far more fundamental. For the foreseeable future, then, microscopy and MRI will remain complementary methods.
Â
To find connectopathies, we will use the methods I outlined above to map reduced connectomes of abnormal and normal brains, and compare them. Some differences may be detectable by dMRI, but subtle ones will require microscopy. We will also compare neuronal connectomes of small chunks of brain using electron microscopy. The use of microscopy poses difficulties, as it must be carried out on the brains of the deceased.
People do bequeath their brains to scienceâthere is a long tradition of such generosityâbut even if we have postmortem brains, many of them present special problems.
One alternative is to search for connectopathies in the brains of animals. Such research will also be important for developing therapies, which are often tested first on animals and only later on humans. The legendary French microbiologist Louis Pasteur produced the first vaccine for rabies by growing the virus in rabbits and then weakening it. The vaccine was tested on dogs before its dramatic first human trial on a nine-year-old boy who had been bitten by a rabid dog.
Studying human mental disorders with animals is no easy task. The rabies virus leads to the same disease, whether it infects rabbits, dogs, or humans. But is there such a thing as an autistic or schizophrenic animal? It's not clear whether such animals occur naturally, but researchers are now attempting to create them using the methods of genetic engineering. Researchers insert the faulty genes associated with autism and schizophrenia into the genomes of animals,
usually mice, with the expectation of giving them analogous disorders. Ideally, such animals would serve as “models” for human disorders, approximations to the real thing.
But this strategy, a variation on Pasteur's, sometimes fails even for infectious diseases. Human immunodeficiency virus (HIV), which causes AIDS in humans, fails to infect many primates, making it difficult to test HIV vaccines. In monkeys, AIDS is caused by simian immunodeficiency virus
(SIV), which is related to HIV but not identical. The lack of a good animal model for human AIDS has slowed down research on finding a cure. Likewise, inserting faulty human genes into animals might not give them autism and schizophrenia. Some analogous but different genetic defect might be necessary.
Because of these uncertainties, the problem of validating animal models for mental disorders has risen to the fore. It's not clear what criteria should be used. Some emphasize similarity of symptoms, but even for infectious diseases this criterion doesn't always work. Sometimes the same microbe can infect both animals and humans but produces very different symptoms. An animal might tolerate infection with little adverse effect at all. And if human genes for autism or schizophrenia turned out to produce very different symptoms in mice, it wouldn't necessarily mean that the mouse models were useless. (Some might argue that it's pointless to compare symptoms, as mental disorders involve behaviors that seem uniquely human.)