Connectome (30 page)
Authors: Sebastian Seung

Â
Figure 46. Cartoon depicting the spikes of three neurons in area HVC of the zebra
Â
As Beethoven booms from the stereo system, the loudspeaker vibrates while electrical signals in the stereo fluctuate wildly. Unlike the fleeting signals, the compact disc remains serene and unchanging. Underneath its label, the plastic surface contains hundreds of millions of microscopic indentations, which encode music as bits of digital information. The plastic will maintain its shape for decades, as the manufacturer guarantees; that stability is why the compact disc will reproduce Beethoven over and over again. Its
material structure
enables it to retain a “memory” of Beethoven's music.
I've compared the spikes of HVC neurons to electrical signals in your compact disc player. Now I'd like to take the analogy further and propose that the HVC connectome is like the compact disc. Let's suppose that it contains a synaptic chain, which no longer changes once a song has crystallized in the adult male. According to this proposal, the HVC connectome retains the memory of the song. Whenever the bird sings, the memory is recalled by converting it into sequential spiking. These signals are fleeting; the material structure of the connections in HVC,
however, remains unchanged.
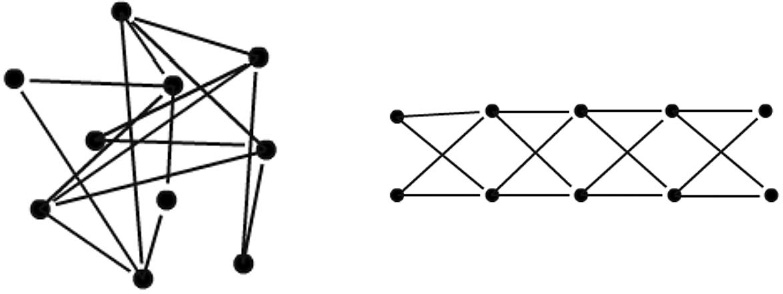
HVC is just a fraction of a cubic millimeter in volume. It should be technically feasible to find its connectome in the near future. Then we could simply examine the connectome to find out whether it's organized like a synaptic chain. This will require some analysis, because it's not obvious whether a connectome contains a chain unless the sequential ordering of the neurons is known. To see why, consider the diagrams shown in Figure 47, both of which have exactly the same connectivity. The neurons on the left have been scrambled to hide the chain. To reveal it,
we must unscramble the neurons to yield the diagram on the right. You can try doing this by hand for our small made-up connectome. But a real HVC connectome is complex enough that a computer would be necessary.
Â
 Â
Â
Â
Figure 47. Synaptic chains, scrambled
(left)
and unscrambled
(right)
Â
Suppose we succeeded in unscrambling the HVC connectome. From the resulting chain we'd be able to guess the order in which the neurons spike during the song. This would amount to reading the memory of song, in the sense that we could guess the activity sequence that is replayed in HVC when the bird sings.
How can we confirm that our reading is correct? Ventris and Chadwick convinced the world they had decoded Linear B because their reading of the clay tablets made sense. If they had failed, the deciphered text would presumably have been gibberish. A stronger test than internal consistency would be to observe and speak with the people who wrote the tablets, but the impossibility of time travel prevents us from doing so.
Similarly, if unscrambling the HVC connectome revealed a synaptic chain, we could already be confident about our reading. Unlike Ventris and Chadwick, we could obtain more conclusive proof without resorting to time travel. Suppose another neuroscientist measures the spike times of HVC neurons during singing but withholds them in order to quiz us. We find the HVC connectome and then read it to guess the spike times. Our examiner compares our guesses with the real spike times; if they match, our reading of the connectome is correct.
To measure the spike times of HVC neurons, our examiner could obtain help from chemists, who have invented ways of staining neurons so that they resemble blinking lights
in the microscope, glowing when they spike and darkening when they fall silent. The images from our examiner's light microscope would also tell us the exact locations of the cell bodies of the HVC neurons. Later on, these locations could be matched up with cell bodies in electron microscopic images of the dead brain. Establishing this correspondence would enable our examiner to compare the real spike times of HVC neurons with the times guessed by reading the connectome.
Of course, it's always possible that we will fail to unscramble the HVC connectome. We might not be able to order
the neurons so that the synapses respect the sequential rule of connection. In other words, no matter how we arrange the neurons in a sequence, there are many connections that go backward or jump
too far ahead. This would mean that the HVC connectome is not organized like a synaptic chain. Such failure would still be progress. For the purpose of advancing science, rejecting models is as important as confirming them.
Â
If the HVC connectome does turn out to be organized like a chain, that would be evidence that it helps retain the bird's memory of its song. But how do memories like this get stored in the first place? Some theoretical neuroscientists have proposed that HVC neurons in young males
are initially driven by random input from some other source. This activates the neurons in random sequences, some of which become reinforced by Hebbian strengthening of connections. These select sequences start to occur more often, thus becoming further reinforced. Ultimately a single sequence is reinforced so much that it dominates all others. This sequence corresponds to the final synaptic chain that we suspect exists in adult males.
According to this proposal, reweighting stores the memory of song. Synapses change in strength, but they are not created or eliminated. The unweighted connectome, which omits information about synaptic strengths, does not contain any of the information in the memory. There is no chance of reading out the spike times of neurons from it. Only the weighted connectome is readable, as only the strong synapses are organized in a chain. In other words, connectomes must include synaptic strengths if they are to be decoded. In principle, that's no problem for connectomics. It should be possible to estimate the strengths of synapses from their appearance in electron microscopic images. As I mentioned earlier, synapses are thought to grow bigger when they get stronger, so size is correlated with strength. Future research should be able to tell us the accuracy of this method for estimating synaptic strength.
Another possibility is that reconnection also plays a role
in the storage of song memory. Maybe synapses not involved in the synaptic chain weaken as the bird learns, and are eventually eliminated. If reconnection does play a role, then even the unweighted connectome might be readable. By attempting to read both unweighted and weighted versions of the HVC connectome, we could conceivably distinguish between the pure reweighting theory of memory and the reweighting-plus-reconnection theory.
Neuroscientists have hypothesized that the other two R's of connectome changeârewiring and regenerationâalso play a role in memory storage, but there is little empirical evidence one way or the other. Fernando Nottebohm and his collaborators have studied regeneration in the brains of canaries and other songbirds. They have shown that HVC shrinks because neurons are eliminated during the part of the year when canaries don't sing. When the singing season comes around again, HVC expands by creating new neurons. Nottebohm's research on regeneration played an important historical role in reviving neuroscientists' interest in the subject, but the function of regeneration remains unclear.
This question could be investigated in a number of interesting ways if the synaptic chain model of HVC is correct. During the off season, does a dormant synaptic chain continue to store the memory of song? When the new neurons enter HVC, do they integrate into the chain? If so, how do they do it? Neural Darwinism predicts that newly created neurons are randomly connected with other neurons. This prediction could be tested empirically by connectomics, with the aid of special stains that mark new neurons.
Similar questions can be asked about the elimination of neurons. What causes neurons to commit suicide? Is it triggered by elimination of synapses and branches, which in turn happens because the neuron fails to integrate into the chain? This hypothesis could be probed using connectomics, through snapshots of neurons caught during the process of dying. To prepare for the off season, are neurons eliminated in such a way that prevents the chain from breaking?
Because of technical limitations, neuroscientists have had to settle for counting increases and decreases in the number of neurons. These studies suggest that regeneration is important, but they do not reveal its exact role in memory. To make further progress, it's crucial to know how new neurons get wired up to the existing organization, and whether the elimination of neurons depends on how they are wired. This kind of information can be provided by connectomics. The function of rewiring could also be studied in HVC by investigating how the growth and retraction of branches of neurons depends on their connections with other neurons.
Â
I've outlined a plan for finding synaptic chains in the HVC connectome and cell assemblies in the CA3 connectome. I've called this “reading memories” from a connectome. More precisely, I've proposed a way of analyzing connectomes to guess activity patterns that are replayed during recollection of a memory. But let me emphasize: That's not the same as knowing what the memory
means.
By analyzing the HVC or CA3 connectomes, we won't know what the bird's song sounds like, or what's in the videos that were previously seen by a human research subject. We might call this the reading of an “ungrounded” memory, one that is divorced from its meaning in the real world.
I already proposed one way of grounding the memory, which is to measure HVC activity in birds as they sing, or CA3 activity in humans while they describe what they're experiencing. Then each neuron could be placed in correspondence with a particular movement or reported idea. This sort of approach uses measurements of spiking in a live brain to ground memories read out after the brain is dead. It's the only approach possible in the near future, as long as we can find only partial connectomes from small chunks of brain.
In the long term, though, I expect that we will be able to find connectomes of entire dead brains. Then it may become possible to ground memories without measuring spiking in live brains. To do this, we'd have to figure out, for example, whether a CA3 neuron is selectively activated by Jennifer Aniston or some other stimulus. Could this be possible by analyzing the pathways that bring information from the sense organs to the CA3 neuron?
It might be, if we employ the hypothesized rules of connection for perceptual neuronsâfor example, “A neuron that detects a whole receives excitatory synapses from neurons that detect its parts.” The Jennifer Aniston neuron might receive inputs from a “blue-eye neuron,” a “blond-hair neuron,” and so on.
For now, researchers are starting to test this partâwhole rule by combining measurements of spiking with connectomics in animals. The first step is to determine the functions of neurons in perception by measuring their spiking in response to various kinds of stimuli, as in the Jennifer Aniston experiment. This is done as described earlier, by staining the neurons so that they blink when active, and observing the neurons through a light microscope. Then researchers image this particular chunk of the brain using an electron microscope to discover how the neurons are connected. Kevin Briggman
and Moritz Helmstaedter have accomplished this feat with retinal neurons, working with Winfried Denk. Studies of neurons in the primary visual cortex have been performed by Davi Bock,
Clay Reid, and their collaborators. This approach, as it develops, will make it possible to see whether there are in fact connections between neurons that detect parts and wholes.
In the coming years the partâwhole rule of connection will be tested in this way. For the sake of discussion, let's suppose that the rule is true, and speculate about how we could use it to read connectomes. The driving idea behind the rule is that a neuron stands on the shoulders of other neurons. We could start by applying the rule to the neurons near the bottom of the hierarchy and guess which stimuli they used to detect. These are the neurons just one step away from the sense organs. Then we could move step by step up the hierarchy, each time guessing the stimuli that neurons detect from the partâwhole rule. Eventually we might reach the top of the heapâCA3 neuronsâand guess which stimuli used to activate them in the live brain. (A neuron that receives connections from neurons that detect floppy ears, sad brown eyes, wagging tail, and loud barkâthat's the neuron that detected your great-great-grandma's dog.
)
Reading memories from dead human brains might sound coolâyou could certainly imagine an entertaining movie being built around this plot deviceâbut it's too far off to be considered seriously as an important practical application of connectomics. What I'm proposing instead as a basic research challenge is to decode the HVC connectome. It would be a way of improving our understanding of how the brain's function depends on the connections between its neurons.  Â
Â
I've discussed several ways of analyzing connectomes: carving them into brain regions, carving them into neuron types, and reading memories from them. These approaches may seem quite different, but all can actually be viewed as the formulation of rules of connection
governing neurons. Each approach in the list is progressively more accurate at predicting connections, because its rules are based on more specific neuronal properties.