Gödel, Escher, Bach: An Eternal Golden Braid (96 page)
Read Gödel, Escher, Bach: An Eternal Golden Braid Online
Authors: Douglas R. Hofstadter
Tags: #Computers, #Art, #Classical, #Symmetry, #Bach; Johann Sebastian, #Individual Artists, #Science, #Science & Technology, #Philosophy, #General, #Metamathematics, #Intelligence (AI) & Semantics, #G'odel; Kurt, #Music, #Logic, #Biography & Autobiography, #Mathematics, #Genres & Styles, #Artificial Intelligence, #Escher; M. C

Incidentally, you may be wondering about this question: "What, according to the Central Dogmap, is Godel's Incompleteness Theorem itself mapped onto?" This is a good question to think about before reading ahead.
The Central Dogmap and the Contracrostipuntus
It turns out that the central dogmap is quite similar to the mapping that was laid out in Chapter IV between the Contracrostipunctus and Godel’s Theorem. One can therefore drew parallels between all three systems.
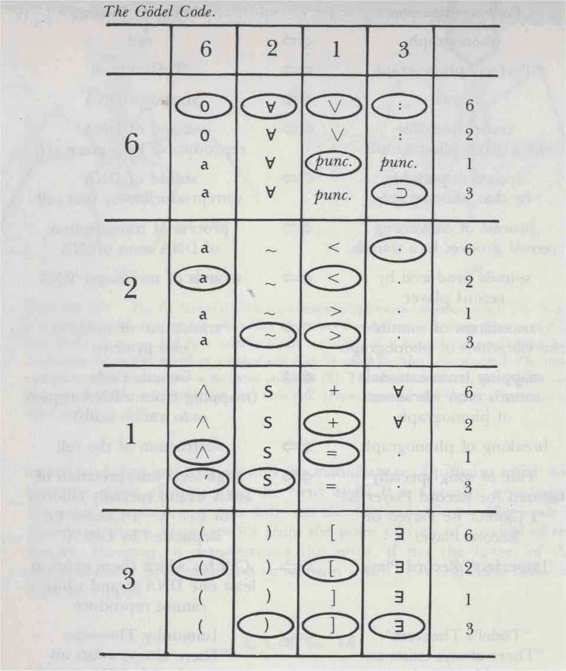
FIGURE 100. The Godel Code. Under this Godel-numbering scheme, each T V7- s'mbol gets one or more codons. The small ovals show how this table subsumes the earlier Godelnumhering table of Chapter IX.
(1) formal systems and strings
(2) cells and strands of DNA
(3) record players and records
In the following chart, the mapping between systems 2 and 3 is explained carefully

The analogue of Godel's Theorem is seen to be a peculiar fact, probably little useful to molecular biologists (to whom it is likely quite obvious):
It is always possible to design a strand of DNA which, if injected into a cell, would, upon being transcribed, cause such proteins to be manufactured as would destroy the cell (or the DNA), and thus result in the non-reproduction of that DNA This conjures tip a somewhat droll scenario, at least if taken in light of evolution: an invading species of virus enters a cell by some surreptitious
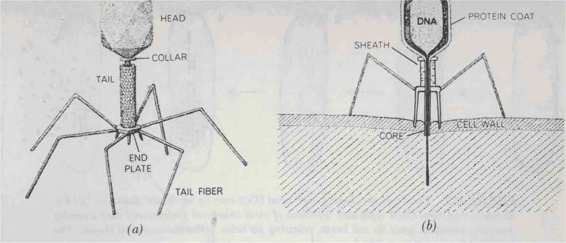
FIGURE 101. The T4 bacterial virus is an assembly of protein components (a). The "head" is a protein membrane, shaped like a kind of prolate irosahedron with thirty facets and filled with DNA. It is attached by a neck to a tail consisting of a hollow core surrounded by a contractile sheathh and based on a spiked end plate to which six fibers are attached. The spikes and fibers affix the virus to a bacterial cell wall (h). The sheath contracts, driving the core through the wall, and viral DNA enters the cell. [From Hanawalt and Haynes, The Chemical Basis of Life, p. 230.1
means, and then carefully ensures the manufacture of proteins which will have the effect of destroying the virus itself! It is a sort of suicide-or Epimenides sentence, if you will-on the molecular level. Obviously it would not prove advantageous from the point of view of survival of the species. However, it demonstrates the spirit, if not the letter, of the mechanisms of protection and subversion which cells and their invaders have developed.
E. Coli vs. T4
Let us consider the biologists' favorite cell, that of the bacterium Escherichia coli (no relation to M. C. Escher), and one of their favorite invaders of that cell: the sinister and eerie T4
phage, pictures of which you can see in Figure 101. (Incidentally, the words "phage" and
"virus" are synonymous and mean "attacker of bacterial cells".) The weird tidbit looks like a little like a cross between a LEM (Lunar Excursion Module) and a mosquito-and it is much more sinister than the latter. It has a "head" wherein is stored all its "knowledge"-namely its DNA; and it has six "legs" wherewith to fasten itself to the cell it has chosen to invade; and it has a "stinging tube" (more properly called its "tail") like a mosquito. The major difference is that unlike a mosquito, which uses its stinger for sucking blood, the T4 phage uses its stinger for injecting its hereditary substance into the cell against the will of its victim. Thus the phage commits "rape" on a tiny scale.
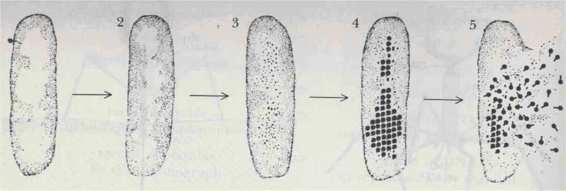
FIGURE 102. Viral infection begins when viral DNA enters a bacterium. Bacterial DNA is disrupted and viral DNA replicated. Synthesis of viral structural proteins and their assembly into virus continues until the cell bursts, releasing particles. [From Hanawalt and Haynes, The Chemical Basis of Life, p. 230.]
A Molecular Trojan Horse
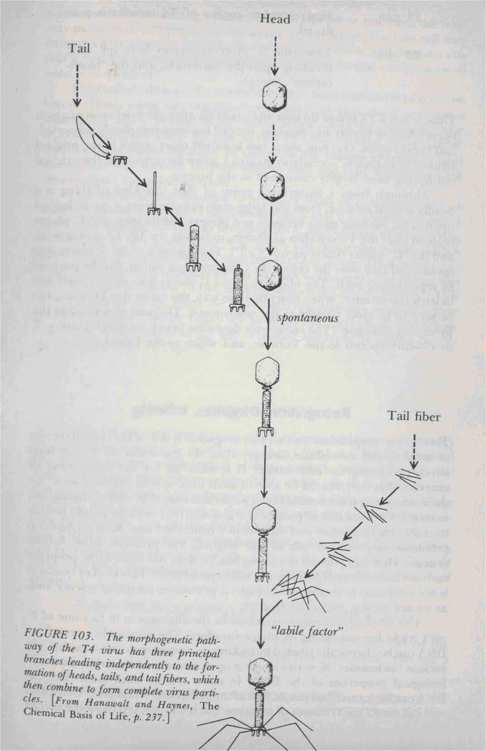
What actually happens when the viral DNA enters a cell? The virus "hopes", to speak anthropomorphically, that its DNA will get exactly the same treatment as the DNA of the host cell. This would mean getting transcribed and translated, thus allowing it to direct the synthesis of its own special proteins, alien to the host cell, which will then begin to do their thing. This amounts to secretly transporting alien proteins "in code" (viz., the Genetic Code) into the cell, and then "decoding" (viz., producing) them. In a way this resembles the story of the Trojan horse, according to which hundreds of soldiers were sneaked into Troy inside a harmless seeming giant wooden horse; but once inside the city, they broke loose and captured it. The alien proteins, once they have been "decoded" (synthesized) from their carrier DNA, now jump into action. The sequence of actions directed by the T4 phage has been carefully studied, and is more or less as follows (see also Figs. 102 and 103): Time elapsed Action taking place
0 min.
Injection of viral DNA.
1 min.
Breakdown of host DNA. Cessation of production of native proteins and initiation of production of alien (T4) proteins. Among the earliest produced proteins are those which direct the replication of the alien (T4) DNA.
5 min.
Replication of viral DNA begins.
8 min.
Initiation of production of structural proteins which will form the "bodies" of new phages.
13 min.
First complete replica of T4 invader is produced.
25 min.
Lysozyme (a protein) attacks host cell wall. breaking open the bacterium, and the "bicentuplets" emerge.
Thus, when a T4 phage invades an E. coli cell, after the brief span of about twenty-four or twenty-five minutes, the cell has been completely subverted, and breaks open. Out pop about two hundred exact copies of the original virus-"bicentuplets"-ready to go attack more bacterial cells, the original cell having been largely consumed in the process.
Although from a bacterium's point of view this kind of thing is a deadly serious menace, from our large-scale vantage point it can be looked upon as an amusing game between two players: the invader, or "T" player (named after the T-even class of phages, including the T2, T4, and others), and the "C" player (standing for "Cell"). The objective of the T player is to invade and take over the cell of the C player from within, for the purpose of reproducing itself. The objective of the C player is to protect itself and destroy the invader.
When described this way, the molecular TC-game can be seen to be quite parallel to the macroscopic TC-game described in the preceding Dialogue. (The reader can doubtless figure out which player-T or C-corresponds to the Tortoise, and which to the Crab.)
Recognition, Disguises, Labeling
.
This "game" emphasizes the fact that recognition is one of the central themes of cellular and subcellular biology. How do molecules (or higher-level structures) recognize each other? It is essential for the functioning of enzymes that they should be able to latch onto special
"binding sites" on their substrates; it is essential that a bacterium should be able to distinguish its own DNA from that of phages; it is essential that two cells should be able to recognize each other and interact in a controlled way. Such recognition problems may remind you of the original, key problem about formal systems: How can you tell if a string has, or does not have, some property such as theoremhood? Is there a decision procedure? This kind of question is not restricted to mathematical logic: it permeates computer science and, as we are seeing, molecular biology.
The labeling technique described in the Dialogue is in fact one of E. colt's tricks for outwitting its phage invaders. The idea is that strands of DNA can be chemically labeled by tacking on a small molecule-methyl-to various nucleotides. Now this labeling operation does not change the usual biological properties of the DNA; in other words, methylated (labeled) DNA can be transcribed just as well as unmethylated (unlabeled) DNA, and so it can direct the synthesis of proteins. But if the host cell has some special
mechanisms for examining whether DNA is labeled or not, then the label may make all the difference in the world. In particular, the host cell may have an enzyme system which looks for unlabeled DNA, and destroys any that it finds by unmercifully chopping it to pieces. In that case, woe to all unlabeled invaders!
The methyl labels on the nucleotides have been compared to serifs on letters. Thus, using this metaphor, we could say that the E. colt cell is looking for DNA written in its
"home script", with its own particular typeface-and will chop up any strand of DNA written in an "alien" typeface. One counterstrategy, of course, is for phages to learn to label themselves, and thereby become able to fool the cells which they are invading into reproducing them.
This TC-battle can continue to arbitrary levels of complexity, but we shall not pursue it further. The essential fact is that it is a battle between a host which is trying to reject all invading DNA, and a phage which is trying to infiltrate its DNA into some host which will transcribe it into mRNA (after which its reproduction is guaranteed). Any phage DNA which succeeds in getting itself reproduced this way can be thought of as having this high-level interpretation: "I Can Be Reproduced in'Cells of Type X". This is to be distinguished from the evolutionarily pointless kind of phage mentioned earlier, which codes for proteins that destroy it, and whose high-level interpretation is the self-defeating sentence: "I Cannot Be Reproduced in Cells of Type X".