Harnessed: How Language and Music Mimicked Nature and Transformed Ape to Man (26 page)
Read Harnessed: How Language and Music Mimicked Nature and Transformed Ape to Man Online
Authors: Mark Changizi
Tags: #Non-Fiction

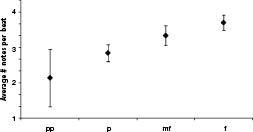
Figure 43
. Data from 60 pieces in Denes Agay’s
An Anthology of Piano Music, Vol. II: The Classical Period
showing that louder portions of music tend to be packed with more notes. Each of 234 contiguous segments of constant loudness were sampled, counting the total number of notes and beats; averages are over these 234 segments. Data collected and analyzed by Caitlin Morris. (Standard errors shown.)
5
Home Pitch
We have discussed how melody consists of one pitch at a time (in the Chapter 4 section titled “Only One Finger at a Time”), which is exactly what we expect if melody has its origins in Doppler shifts. We also mentioned earlier in that chapter (in the section titled “Why Pitch Seems Spatial”) that melodic pitch tends to change in a fairly continuous fashion. At any one time, then, melody is at one pitch, and changes pitch roughly as if it is “moving” through pitches. Thinking of melody as if it is an unknown creature we wish to better understand, it is natural to ask about melody’s home and where it roams within the space of pitches. In this Encore section we’ll discuss three facets of melody’s home range and foraging behavior.
Let’s begin with one of the most salient features about the Doppler effect, which is that for a mover at constant speed there is a maximum and minimum pitch the mover can attain, these occurring when the mover is headed directly toward and directly away from the listener, respectively. Doppler pitches for any mover are therefore bound to a fixed home range. If Doppler pitches are confined in this sense to a fixed home range, then the music-is-movement theory predicts that melodies, too, should tend to confine themselves to a fixed home range. Melodies should tend to behave as if there is an upper or lower boundary to pitch.
Does
melody move around as if bound within an invisible fence, as predicted, or does melody move more freely? Although melody is highly variable, it has long been noticed that any given melody tends to confine itself to a fairly fixed window of pitches called its tessitura. The notion of tessitura allows that the melody may occasionally punch through a barrier, but the barriers are still worthy of recognition because of their tendency to hold the pitch inside. The tessitura is, I submit, music’s implicit recognition that a single constant-speed mover has a fixed range of Doppler pitches it can express.
Melody, then, has a fixed home range, consistent with Doppler shifts. Let’s now look into how melody spends its time within its home range. If melody really is acting like Doppler shifts, then melody should distribute itself within its home range in a similar manner to Doppler pitches. How
do
Doppler pitches distribute themselves within their home range? In particular, for a mover in your vicinity carrying out behaviors, in which directions does the mover tend to go? Many of the movers around you are just doing their own thing, carrying out actions that do not involve the fact that you are there. These movers will tend to go in any direction relative to you. But even movers who are interacting in some way with you will tend not to strongly favor some directions over others. For example, performers on a stage will, over the course of the show, move in all directions relative to any audience member. In fact, although their actions onstage may be highly intricate, they will often be very roughly summarized as moving in circles out in front of the listener, an illustrative case we had used earlier in Figure 25 of Chapter 4. Such circle-like behavior tends to sample broadly from all directions. Individual short bouts of behavior, then, can be anywhere in the Doppler pitch range.
Across
bouts of behavior, then, Doppler pitches tend to occur with fairly uniform probability over the Doppler range. For melody, then, we expect that individual melodic themes will be highly variable in their pitch distribution, but we also expect that, on average, these themes will sample pitches within their tessitura fairly uniformly.
That is, in fact, what we found among the classical themes. Despite wide variability from theme to theme, across the 10,000 classical themes the average distribution of notes across the tessitura is fairly flat, as shown in Figure 44. One might have expected to find that, say, melody strongly favors a single central pitch, and meanders away from this pitch as if tied to it by an elastic band, in which case the expected distribution would be disproportionately found on or near that pitch, and would fall steeply lower and lower the farther away a pitch is from that central one. Melody does not, however, behave like this. Melody is more Doppler-like, sampling pitches within its home range in a fairly egalitarian fashion.
Both Doppler and melodic pitches have a fixed home range, and both tend to roam fairly uniformly over their home. Let’s now ask whether some regions within the home range are “stickier.” That is, are there regions of the tessitura where, if the melody goes there, it takes longer to get out? This differs from what we just finished discussing—that concerned the
total
amount of time (actually, the total number of notes) spent in regions of the tessitura, whereas we are now asking how long in duration each singular visit to a region tends to be. To understand sticky pitches in melody, we look to the Doppler shifts of movers and ask whether any Doppler pitches are sticky.
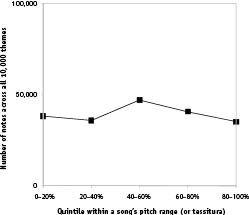
Figure 44
. Distribution showing where within a theme’s tessitura notes tend to lie, among the nearly 200,000 notes in the 10,000 classical themes. (That is, for each note, its position within its theme’s tessitura was determined and its quintile recorded. The plot shows the distribution of these values.) One can see that themes tend to have pitches sampling widely across the tessitura, with little tendency to favor some parts of the tessitura over others. (The shape looks identical if the distribution for each theme is separately determined, and all 10,000 distributions averaged, and the error bars on such a plot are far too small to discern.) Note again that this analysis uses the tessitura for each piece, rather than measuring the number of semitones away from the average pitch in the song; the latter analysis would lead to a more normal distribution, falling quickly in probability away from the average, something researchers Tierney, Russo, and Patel found in 2008. In light of the result here, the normal distribution they found is due to the distribution of tessitura widths, not the distribution of pitches within the tessitura.
There are, indeed, sticky Doppler pitches; they are the pitches near the top and bottom of the pitch range. To see why, imagine again a mover who is running in circles out in front of you, as depicted in Figure 45. Even though the mover is going through all directions uniformly, the pitches tend to change most quickly when the mover is whizzing horizontally by, either dropping quickly in pitch when passing nearby, or rising quickly in pitch when whizzing by at the far side of the circular path. When the mover is in the approach or withdrawal parts of the path, on the other hand, the pitch is fairly stable and high or low, respectively. Figure 45 shows these four segments of the circular path of the mover, and one can see that the pitch in the “toward” and “away” segments is much more stable than in the two “whiz by” segments. Doppler pitches vary less quickly near the top and bottom of their home range. The prediction, then, is that melodic pitch tends to change more slowly near the top and bottom of the tessitura. Does it? To test this, Sean Barnett measured the durations for all notes among the 10,000 classical themes. Each note was classified as a bottom, intermediate, or top note for its theme, and the average duration was computed for each of the three categories. Figure 46 shows these average durations, and one can see that lowest and highest notes in themes tend to be 17 percent longer in duration than notes with intermediate pitches.
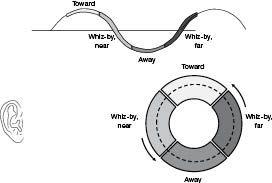
Figure 45
. Doppler pitch changes slowly when near the maximum and minimum of the Doppler pitch range. Melodies also share this, as shown in Figure 46.

Figure 46
. Across the 10,000 classical themes, this plot shows the average duration of the minimum, intermediate, and maximum pitch in a theme. One can see that the minimum and maximum pitch in a theme each tend to be longer in duration than intermediate pitches. (Averages of durations were computed in logarithmic space. Error bars are too small to see.)
In this section we looked at three facets of melody’s home range. We saw that (i) melodies typically
have
a fixed home range, called the tessitura; (ii) melodies tend to distribute themselves fairly uniformly within their home range; and (iii) melodies tend to dwell longer at the edges of their range. Melody behaves in these ways, I am suggesting, because Doppler shifts behave in these ways. Melody is broadly Doppler-like in the home it keeps and the manner in which it distributes its movements and time throughout its home.
6
Fast Tempo, Wide Pitch
In the previous section we examined melody’s home range—its size, and melody’s hangouts within it. One facet of melody we discussed was that it tends to remain in a cage, called the tessitura, and I am suggesting that the top and bottom of the tessitura correspond to the Doppler pitches when the fictional mover is directed toward and away from you, respectively. But remember that Doppler shifts are greater when the mover has greater speed. A car driving past you at a crawl will have a small difference between its high approaching pitch and its low moving-away pitch. But if you stand at the side of the freeway, the difference in pitch as the cars pass you will be much greater. It follows from these simple observations that faster-tempo pieces of music should have bigger home ranges for their melodies. That is, if melodic contour has been culturally selected to mimic the Doppler shifts of movers, then the prediction is that music with a faster tempo (more beats per minute) should have a wider tessitura.
To test this, Sean Barnett and I measured the tempo and tessitura width of the melodies of all the pieces in the
Classical Fake Book
(Hal Leonard Corp.). (We did not use the
Dictionary of Musical Themes
here because it does not include tempo data.) Figure 47 shows how tessitura width varies with tempo (for just those pieces originally intended for keyboard). One can see that although tessitura width does not change for the several low tempos, it rises among the faster tempos. Tessitura width increases with greater tempo, as predicted from the fact that the Doppler pitch home range widens as mover speed increases. This is particularly striking because themes with wider tessituras tend to be more difficult to play, and so one might predict that wider-tessitura music would go with a
slower
tempo, but this is the opposite of what we in fact find.
One might wonder whether this result could be due, instead, to a general phenomenon in which faster-tempo music tends simply to amplify musical qualities, whatever they may be. Caitlin Morris measured the range of loudness levels—the “loudness-tessitura” width—and the tempo for a sample of 55 pieces in Denes Agay’s piano anthology,
The Classical Period.
Figure 48 shows how the width of the loudness range varies with tempo, and one can see that there is no trend. The pitch tessitura width does not, then, increase in Figure 47 merely because of some general proclivity to amplify musical qualities at higher tempos. In fact, the
lack
of change in “loudness-tessitura” width as a function of tempo is something the music-is-movement theory
does
predict, assuming that loudness in music is primarily driven by proximity, as we discussed in detail in the “Nearness versus Stompiness” section of Chapter 4. Imagine that a mover carries out a bout of behavior in your vicinity at low speed. Now imagine this mover is asked to repeat the same bout of behavior, but this time moving much more quickly—that is, at a higher tempo. In each case the mover is, we presume, going through the same sequence of spatial coordinates, and thus the same sequence of distances from the listener. And so it immediately follows that the mover courses through the same sequence of loudnesses no matter whether moving slowly or quickly. The music-is-movement theory predicts, then, that, unlike pitch, the range of loudnesses should
not
change as a function of the music’s tempo—faster music,
same
loudness range—and that’s what we found.
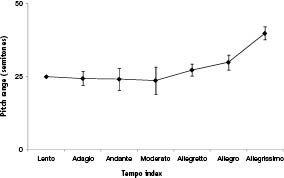
Figure 47
. Tessitura (of melody) width versus tempo, among all 92 pieces for keyboard in the
Classical Fake Book
for which tempo data could be acquired. One can see that faster music tends to have wider tessituras, consistent with the Doppler interpretation of melodic pitch. (We found the same result when we used the data for all pieces.) The
Classical Fake Book
was used for two reasons. First, it is helpful because fake books are not cluttered with the notes from the chords (chords are notated via letter labels). Second, it is the only classical fake book I possess, so it amounted to an easy-to-get, unbiased sample.