Junk DNA: A Journey Through the Dark Matter of the Genome (12 page)
Read Junk DNA: A Journey Through the Dark Matter of the Genome Online
Authors: Nessa Carey

This is falling into the surprisingly common trap of focusing on what we see, rather than what we don’t see. The reason that we see babies born with trisomies of chromosomes 13, 18 and 21 is because these are relatively benign, unlikely though that sounds. These are three of the smallest chromosomes and they each contain relatively few genes. Generally, the larger the chromosome, the greater the number of genes it contains. So the reason we never see trisomy of chromosome 1, for example, is because of its size. Chromosome 1 is very large and contains a lot of genes. If an egg and sperm fuse and create a zygote with three copies of this chromosome, there will be overexpression of such a large number of genes that the cell function will be disrupted catastrophically, leading to extremely early destruction of the embryo. This probably occurs before the woman is even aware she is pregnant.
For women aged between 25 and 40, the success rates for in vitro fertilisation using donated eggs are not affected by age.
31
But the likelihood of a woman becoming pregnant naturally does decline after her mid-20s. The difference between these two situations suggests that the mother’s age critically affects her eggs, rather than her uterus. We already know from Down’s Syndrome that maternal age influences the success of chromosome segregation into the eggs. So it’s not too big a leap to hypothesise that the decline in pregnancy rates after the mid-20s may be in part due to very early failures of embryo development, as a result of malfunctioning centromere activity and the creation of eggs with disastrous misallocation of large chromosomes.
Footnotes
a
Specifically, budding yeast such as
Saccharomyces cerevisiae
.
b
Caenorhabditis elegans
.
c
This unit of 171 base pairs is called an alpha (α) satellite repeat.
d
These are called histone H3.
7. Painting with Junk
In a twelve-month period from 2011 to 2012, 813,200 babies were born in the UK.
1
Using the rates quoted in the previous chapter, we can estimate that nearly 1,200 of these babies had Down’s Syndrome, around 270 had Edward’s Syndrome and just under 120 had Patau’s Sydrome. That’s a very small number of cots in a nursery of over three-quarters of a million babies. This is consistent with the concept that having too many copies of a chromosome is very damaging: in general we would not expect high survival rates when it occurs.
Which makes it all the more surprising to learn that about half of the babies born in that period – that’s over 400,000 children – were born with one chromosome too many. Yes, one in two of us. Even more confusingly, the extra chromosome isn’t some tiny little genetic remnant. It’s a really big chromosome. How on earth can this be, when one extra copy of a very small chromosome can cause devastating conditions such as Edward’s or Patau’s Syndromes?
The culprit here is known as the X chromosome, and it’s prevented from causing harm by a process that relies utterly on junk DNA. But before we move to exploring how this protection happens, we need to explore the nature of the X chromosome itself.
Most of the time the chromosomes in a cell are very long and stringy, and difficult to distinguish from each other. They appear like a great bundle of tangled wool when viewed under a normal light microscope. But when a cell is getting ready to divide, the chromosomes become very structured and compact, and are really discrete entities. If you know the right techniques, you can
isolate all the compacted chromosomes from a nucleus, stain them with specific chemicals and examine the individual ones through a microscope. At this stage they look more like separate skeins of embroidery wool, with the centromere as the little tube of paper that holds the skeins in place.
By analysing photos of the whole complement of chromosomes in a human cell, scientists were able to identify each individual chromosome. They literally used to cut and paste the individual chromosome pictures to arrange them in order. This is how researchers discovered the causes of Down’s, Edward’s and Patau’s Syndromes, by analysing the chromosomes in cells taken from affected children.
But before identifying the underlying problems in these serious conditions, the early researchers discovered the fundamental organisation of our genetic material. They showed that the normal number of chromosomes in a human cell is 46. The exceptions are the eggs and the sperm, which each have 23. Our chromosomes are arranged in pairs, inherited equally from our mother and father. In other words, one copy of chromosome 1 from mum and one from dad. The same for chromosome 2, and for the others.
This is true for chromosome 1 up to chromosome 22. These are known as the autosomes. If we only looked at the autosomes in a cell, we would not be able to tell if the cell was from a female or a male. But this information becomes immediately apparent if we look at the last remaining pair of chromosomes, known as the sex chromosomes. Females have two identical large sex chromosomes, known as X. Males have one X chromosome and a very small chromosome, called Y. These two situations are shown in Figure 7.1.
The Y chromosome may be small, but it has an amazing impact. It’s the presence of the Y chromosome that determines the sex of the developing embryo. It only contains a small number of genes, but these are vitally important in governing gender.
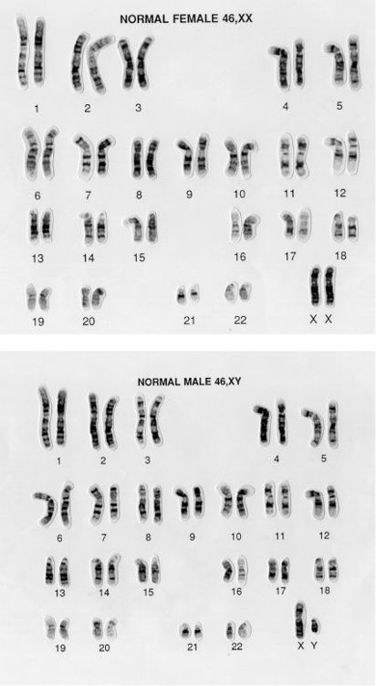
Figure 7.1
Standard female and male karyotypes, showing all the chromosomes present in a cell. The upper panel shows a female karyotype, the lower a male one. The only difference is in the last pair of chromosomes. Females have two large X chromosomes, males have one large X and a small Y. (Wessex Regional Genetics Centre, Wellcome Images)
In fact, this is predominantly controlled by just one gene
a
2
which drives creation of the testes. This in turn leads to production of the hormone testosterone, which results in masculinisation of the embryo. Remarkably, a recent study has shown that just this and one other gene are sufficient not just to create male mice, but also for these mice to generate functional sperm and to father pups.
3
The X chromosome, on the other hand, is very large, containing over 1,000 genes.
4
This creates a potential problem. Males only have one copy of the X chromosome and hence one copy of each of these genes. But females have double that number, so in theory could produce twice as much of the products encoded by the X chromosome as males. The trisomic conditions described in Chapter 6 demonstrated that even a 50 per cent increase in expression of the genes from a small chromosome has a hugely detrimental effect on development. How then can females tolerate a 100 per cent increase in expression of over 1,000 genes, compared with males?
Women have an off switch
The answer is that they don’t. Females produce the same amount of X chromosome-encoded protein expression in their cells as males. They achieve this by a remarkably ingenious arrangement whereby one X chromosome is switched off in every cell. This is known as X-inactivation. Not only is it essential for human life, the process by which it occurs opened up new and totally unanticipated areas of biology that are still the subject of intense scrutiny.
One of the oddest things we have come to realise is that our cells can count the number of X chromosomes. Male cells contain an X and a Y chromosome and they never inactivate the single X. But sometimes males are born who have two X chromosomes and
one Y. They are still males, because it’s the Y chromosome that drives masculinisation. But their cells inactivate the extra X, just as female cells do.
A similar thing happens in females. Sometimes females are born who have three X chromosomes in each cell. When this happens, the cells shut down two X chromosomes instead of one. The flip side of this is when females are born who only have one X chromosome. In this case, the cell doesn’t shut it off at all.
In addition to being able to count, our cells are also able to remember. When a female produces eggs, she usually only gives each egg one of each pair of chromosomes, including the X chromosome. A male produces sperm that contain either an X or a Y chromosome. When a sperm that contains an X chromosome fuses with an egg, the resulting single-cell zygote contains two X chromosomes and both are active. But very early in development, after just a few rounds of cell division, one X chromosome is inactivated in each cell of the embryo. Sometimes it’s the X that came from father, sometimes the X that came from mother. Every daughter cell that subsequently develops switches off the same X chromosome as its parental cell. This means that of the 50 trillion or so cells in the adult female body, on average about half will express the X chromosome that was provided by the egg, and the other half will express the X chromosome that was provided by the sperm.
When an X chromosome is inactivated, it adopts a very unusual physical conformation. The DNA becomes incredibly compacted. Imagine you and a friend each take hold of opposite ends of a towel. You start turning your end of the towel clockwise, and your friend does the same at the other end. Pretty quickly, the towel will start twisting in the middle, and the two of you will be pulled closer together. Now imagine that the towel is about five metres in length, but you manage to keep twisting it until it’s a dense clump of towel only a millimetre in linear length. By this stage, the
towel is extraordinarily tightly wound up. Essentially, the X chromosome becomes as tightly compacted as that towel. One of the consequences is that it forms a dense structure that can be seen when looking at the nucleus of a female cell down a microscope, when all the other chromosomes are long and stringy and can’t be visualised. The condensed X chromosome is called the Barr body.
In order to try to understand how X chromosome inactivation happens, scientists studied unusual cell lines and mouse strains. These focused on examples where parts of the X chromosome had been lost, or where bits of it had been transferred to other chromosomes. Some cells that had lost part of the X chromosome were still able to inactivate one of their X chromosomes, as shown by the presence of the Barr body. But cells that had lost a different part of the X weren’t able to form Barr bodies, showing that they hadn’t inactivated a chromosome.
Where parts of the X chromosome had been transferred to other chromosomes, sometimes these abnormal chromosomes were inactivated, and other times they weren’t. It all depended on which bit of the X chromosome had been transferred.
These data enabled researchers to narrow down the region on the X chromosome that was key for inactivation. Rationally enough, they called this region the X inactivation centre. In 1991, a group reported that this region contained a gene that they called Xist.
b
Only the Xist gene on the inactive chromosome expressed Xist RNA.
5
,
6
This made perfect sense, because X inactivation is an asymmetric process. In a pair of equivalent X chromosomes, one is inactivated and one is not. So it seemed consistent that this process would be driven by a scenario where one chromosome expresses a gene and the other doesn’t.
A very large bit of junk
It was obvious that the next question would be to ask how Xist works and the first thing that researchers did was to try to predict the sequence of the protein that it produced. This is usually relatively straightforward. Once they had found the sequence of the Xist RNA molecule, all that the scientists had to do was run this through a simple computer program that would predict the encoded amino acid sequence. Xist RNA is very long, about 17,000 bases. Each amino acid is encoded by a block of three bases, so a 17,000-base RNA could theoretically code for a protein of over 5,700 amino acids. But when the Xist RNA sequence was examined, the longest run of amino acids was just under 300. This was despite the fact that the Xist RNA was spliced, in the way we first saw in Chapter 2, so had lost all the intervening junk sequences.
The ‘problem’ was that the Xist RNA was liberally scattered with sequences that don’t code for amino acids, but which act as stop signals when protein chains are being built up. We can envisage this as being a little like trying to build a tall tower out of LEGO. It is perfectly straightforward until someone hands you one of those roof bricks that doesn’t have any of the attachment nodes on the top. Once you insert this brick, your tower can’t get any bigger.
If Xist did encode a protein, it would seem very odd that a cell would go to the effort of creating an RNA that was 17,000 bases
c
in length just to produce a protein that could have been encoded by an RNA of about 5 per cent of that length. Researchers in the field realised relatively quickly that this wasn’t what was happening. The reality was much stranger.