In Search of Memory: The Emergence of a New Science of Mind (18 page)
Read In Search of Memory: The Emergence of a New Science of Mind Online
Authors: Eric R. Kandel
Tags: #Psychology, #Cognitive Psychology & Cognition, #Cognitive Psychology

From her systematic studies of H.M., Milner extracted three important principles of the biological basis of complex memory. First, memory is a distinct mental function, clearly separate from other perceptual, motor, and cognitive abilities. Second, short-term memory and long-term memory can be stored separately. Loss of medial temporal lobe structures, particularly loss of the hippocampus, destroys the ability to convert new short-term memory to new long-term memory. Third, Milner showed that at least one type of memory can be traced to specific places in the brain. Loss of brain substance in the medial temporal lobe and the hippocampus profoundly disrupts the ability to lay down new long-term memories, whereas losses in certain other regions of the brain do not affect memory.
Milner thus disproved Lashley’s theory of mass action. It is only in the hippocampus that the various strands of sensory information necessary for forming long-term memory come together. Lashley never went below the surface of the cortex in his experiments. Moreover, Milner’s finding that H.M. had good long-term memory for events that happened prior to the surgery showed clearly that the medial temporal lobe and the hippocampus are not the permanent storage sites of memory that has been in long-term storage for some time.
We now have reason to believe that long-term memory is stored in the cerebral cortex. Moreover, it is stored in the same area of the cerebral cortex that originally processed the information—that is, memories of visual images are stored in various areas of the visual cortex, and memories of tactile experiences are stored in the somatosensory cortex (figure 8–6). That explains why Lashley, who used complex tasks based on several different sensory modalities, could not erase his rats’ memories fully by removing selected sections of the cortex.
For many years Milner thought H.M.’s memory defect was complete, that he was unable to convert any short-term memory to long-term memory. But in 1962 she demonstrated another principle of the biological basis of memory—the existence of more than one kind of memory. Specifically, Milner found that in addition to conscious memory, which requires the hippocampus, there is an unconscious memory that resides outside the hippocampus and the medial temporal lobe. (This distinction had been proposed on behavioral grounds in the 1950s by Jerome Bruner at Harvard, one of the fathers of cognitive psychology.)
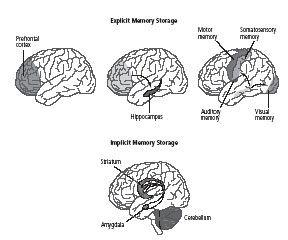
8–6 Explicit and implicit memories are processed and stored in different regions in the brain
. In the short term, explicit memory for people, objects, places, facts, and events is stored in the prefrontal cortex. These memories are converted to long-term memories in the hippocampus and then stored in the parts of the cortex that correspond to the senses involved—that is, in the same areas that originally processed the information. Implicit memories of skills, habits, and conditioning are stored in the cerebellum, striatum, and amygdala.
Milner thus demonstrated this distinction by showing that the two forms of memory require different anatomical systems (figure 8–6). She found that H.M. could learn and remember some things over the long term—that is, he had a kind of long-term memory that does not depend on the medial temporal lobe or the hippocampus. He learned to trace the outline of a star in a mirror and his skill at tracing improved from day to day, just as it would in a person without brain damage (figure 8–7). Yet, even though his performance improved at the beginning of each day’s test, H.M. could never remember having performed the task on an earlier day.
The ability to learn a drawing skill proved to be just one of a number of abilities that were intact in H.M. Moreover, this and other learning abilities described by Milner proved to be remarkably general and applied equally well to other people with damage to the hippocampus and the medial temporal lobe. Thus Milner’s work revealed that we process and store information about the world in two fundamentally different ways (figure 8–6). It also revealed once again that, as with Broca and Wernicke, one learns a great deal from the careful study of clinical cases.
Larry Squire, a neuropsychologist working at the University of California, San Diego, expanded on Milner’s finding. He did parallel experiments on memory storage in humans and in animals. These studies and those of Daniel Schacter, now at Harvard University, described the biology of two major classes of memory.
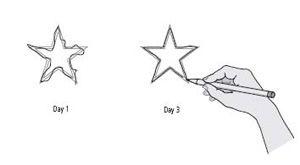
8–7 Despite his obvious loss of memory, H.M. could learn and retain new
skills. During his first attempt on Day 1 (left), H.M. made many errors tracing a star he could see only in a mirror. During his first attempt on Day 3 (right), H.M. retained what he had learned through practice—even though he had no recollection of the task.
What we usually think of as conscious memory we now call, following Squire and Schacter, explicit (or declarative) memory. It is the conscious recall of people, places, objects, facts, and events—the memory that H.M. lacked. Unconscious memory we now call implicit (or procedural) memory. It underlies habituation, sensitization, and classical conditioning, as well as perceptual and motor skills such as riding a bicycle or serving a tennis ball. This is the memory H.M. retained.
Implicit memory is not a single memory system but a collection of processes involving several different brain systems that lie deep within the cerebral cortex (figure 8–6). For example, the association of feelings (such as fear or happiness) with events involves a structure called the amygdala. The formation of new motor (and perhaps cognitive) habits requires the striatum, while learning new motor skills or coordinated activities depends on the cerebellum. In the simplest animals, including invertebrates, implicit memory for habituation, sensitization, and classical conditioning can be stored within the reflex pathways themselves.
Implicit memory often has an automatic quality. It is recalled directly through performance, without any conscious effort or even awareness that we are drawing on memory. Although experiences change perceptual and motor abilities, those experiences are virtually inaccessible to conscious recollection. For example, once you learn to ride a bicycle, you simply do it. You do not consciously direct your body: “Now push with my left foot, now my right….” If we paid that much attention to each movement, we would probably fall off the bicycle. When we speak, we do not consider where in the sentence to place the noun or the verb. We do it automatically, unconsciously. This is the type of reflexive learning studied by the behaviorists, notably Pavlov, Thorndike, and Skinner.
Many learning experiences recruit both explicit and implicit memory. Indeed, constant repetition can transform explicit memory into implicit memory. Learning to ride a bicycle initially involves conscious attention to one’s body and the bicycle, but eventually riding becomes an automatic, unconscious motor activity.
Philosophers and psychologists had already anticipated the distinction between explicit and implicit memory. Hermann Helmholtz, the first person to measure the speed of conduction of the action potential, also worked on visual perception. In 1885 he pointed out that a great deal of mental processing for visual perception and for action occurs on an unconscious level. In 1890, in his classic work
The Principles of Psychology
, William James expanded on this idea, writing separate chapters on habit (unconscious, mechanical, reflexive action) and memory (conscious awareness of the past). In 1949 the British philosopher Gilbert Ryle distinguished between knowing
how
(the knowledge of skills), and knowing
what
(knowledge of facts and events). Indeed, a central premise of Freudian psychoanalytic theory, enunciated in 1900 in
The Interpretation of Dreams
, is an extension of Helmholtz’s idea that experiences are recorded and recalled not only as conscious memories, but also as unconscious memories. Unconscious memories are ordinarily inaccessible to consciousness, but they nevertheless exert powerful effects on behavior.
While Freud’s ideas were interesting and influential, many scientists were not convinced of their truth in the absence of experimental inquiry into how the brain actually stores information. Milner’s star-tracing experiment with H.M. was the first time a scientist had uncovered the biological basis of a psychoanalytic hypothesis. By showing that a person who has no hippocampus (and therefore no ability to store conscious memories) can nonetheless remember an action, she validated Freud’s theory that the majority of our actions are unconscious.
WHENEVER I RETURN TO BRENDA MILNER’S PAPERS ON H.M
., I am impressed yet again by how much these studies clarified our thinking about memory. Pierre Flourens in the nineteenth century and Karl Lashley well into the twentieth century thought of the cerebral cortex as a bowl of porridge, in which all regions were similar in how they worked. For them memory was not a discrete mental process that could be studied in isolation. But when other scientists began to track not only cognitive processes but also various memory processes to distinct regions of the brain, the theory of mass action was dismissed once and for all.
Thus by 1957, having read Milner’s initial paper and having some insight into where memory is stored in the brain, the question of how memory is stored in the brain had become the next meaningful scientific question for me. As I settled into Wade Marshall’s laboratory, I began to think that this question would be an ideal challenge. Moreover, I thought that the answer would best be pursued by examining the cells involved in the storage of specific explicit memories. I would plant my flag midway between my interests in clinical psychoanalysis and the basic biology of nerve cells and proceed to investigate the territory of explicit memory “one cell at a time.”
P
rior to Brenda Milner’s discoveries, many behaviorists and some cognitive psychologists had followed the lead of Freud and Skinner and abandoned biology as a useful guide to the study of learning and memory. They had done so not because they were dualists, like Descartes, but because they thought that biology was unlikely to play a significant role in studies of learning in the near future. Indeed, Lashley’s influential work made it seem that the biology of learning was essentially incomprehensible. In 1950, toward the end of his career, he wrote, “I sometimes feel, in reviewing the evidence on the localization of the memory trace, that the necessary conclusion is that
learning is just not possible
[italics added].”
Milner’s work changed all that. Her discoveries that certain regions of the brain are necessary for some forms of memory provided the first evidence of
where
different memories are processed and stored. But the question of
how
memory is stored remained unanswered, and it fascinated me. Although I had only the most rudimentary preparation for researching how memory is stored in the nervous system, I was eager to give it a try—and the environment at NIH encouraged a certain boldness. All around me research on various problems in the spinal cord, first outlined by Sherrington, was being conducted at the cellular level. Ultimately, cellular studies of memory had to answer a number of key questions: What changes occur in the brain when we learn? Do different types of learning involve different changes? What are the biochemical mechanisms of memory storage? These questions were swirling in my mind, but such questions do not translate easily into useful experiments.
I wanted to begin where Milner had left off. I wanted to tackle the most complex and interesting aspect of memory—the formation of long-term memory for people, places, and things that she found lacking in H.M. I therefore wanted to focus on the hippocampus, which Milner had shown was essential for the formation of new long-term memories. But my ideas about how to tackle the biology of memory in the hippocampus were not only vague, they were naïve.
As a first step, I asked a simple question: Do nerve cells that participate in memory storage have easily recognizable distinguishing features? Were the nerve cells of the hippocampus—cells that are presumably critical for memory storage—physiologically different from motor neurons in the spinal cord, the only other well-studied neurons in the mammalian central nervous system? Possibly, I thought, the properties of hippocampal neurons would reveal something about how memory is recorded.
I was emboldened to try this technically demanding study because Karl Frank, who worked in the lab next door to me, and John Eccles in Australia were using microelectrodes to study individual motor neurons in the spinal cord of cats. Their electrodes were identical to the ones I had used to listen in on crayfish cells. Although Frank himself thought that studying the hippocampus was formidable and risky, he was not discouraging.
Marshall had only one laboratory and two postdoctoral fellows, Jack Brinley and me. Jack had obtained a medical degree from the University of Michigan and had started work on a Ph.D. in biophysics at Johns Hopkins University just before he came to NIH. His proposed thesis was on the movement of potassium ions through the membrane of neurons in the autonomic nervous system. Because Wade liked the cerebral cortex, Jack shifted his focus a bit and studied potassium movement through the cerebral cortex in response to spreading cortical depression, a seizure process that Marshall had been interested in for several years. This was a perfectly good problem, but not one that interested me. Jack felt the same way about the hippocampus. So we worked out a compromise: we would share the lab. He would use it half of the time and I would help him, and I would use it the other half and he would help me.
This arrangement was working well when all of a sudden Marshall thrust upon us a third person, a new postdoctoral fellow, Alden Spencer, who had just graduated from the University of Oregon Medical School. The idea that now the lab would be shared by three independent projects with each of us having even less time in the lab to work on our own studies produced apprehension in both Jack’s heart and mine. We each worked feverishly to persuade Alden to join our particular project.
To my delight, it took little effort to convince Alden that we should work together on the hippocampus. Part of my success, which I did not realize until later, was due to the fact that Alden never for a moment considered working with Jack, whose project required using a radioactive form of potassium. Alden was a bit of a hypochondriac and was frightened to death of radioactivity.
MY RESEARCH TOOK AN EXTREMELY FORTUNATE TURN WITH
Alden’s arrival. Born in Portland, he was a liberal in the best Oregonian tradition of independent thought based on moral rather than narrow political considerations (figure 9–1). Alden’s father, a perpetual student, at once a freethinker and a religious man, was a conscientious objector during World War I and was recruited into the noncombatant corps. After the war, he went to divinity school in British Columbia and served for a while as pastor of a small church. He then went back to school at Stanford University, where he studied mathematics and statistics and later worked as a statistician for the civil service in Oregon.
Alden completely changed my narrow view of life outside the East Coast. He was strongly independent, with an original turn of mind, a great interest in music and art, and an enthusiasm for life that made him exciting to be with. He had novel insights about most things he experienced: a lecture, a concert, a tennis match. His creativity was so abundant and came to him so readily that he was always extending himself to something new, immersing himself in yet another problem. Alden had a considerable musical talent as well, having played the clarinet in the Portland Symphony Orchestra. His wife, Diane, was a fine pianist. Moreover, Alden was extremely modest and expressed these deep creative interests in an utterly unpretentious way. Denise and I soon became good friends with them, and the four of us routinely attended the Library of Congress’s weekly chamber music recitals featuring the renowned Budapest String Quartet.

9–1
Alden Spencer (1931–1977), with whom I had the privilege of collaborating at the NIMH from 1958 to 1960 and who later joined me at NYU Medical School and at Columbia University. Alden made major contributions to the understanding of the hippocampus, the modification of simple reflex responses by learning, and the perception of touch. (From Eric Kandel’s personal collection.)
Among Alden’s many talents were surgical skills, a good knowledge of the anatomical organization of the brain, and insights into what questions were scientifically important. Although he had no experience with intracellular recording, he had done some excellent electrophysiological research on the brain, studying how the pathways between the thalamus and the cortex contribute to the various brain rhythms displayed on EEGs (electroencephalograms). Alden was great company. We talked science incessantly and reinforced each other’s audacity. Once we decided it was important, we were not reluctant to tackle any difficult problem, such as trying to obtain recordings from individual cortical neurons in an intact brain.
Soon after we started we had our first successful experiment. I shall never forget it. I worked all morning and part of the afternoon to complete the surgery that exposed a cat’s hippocampus. Late in the afternoon Alden took over and was advancing the recording electrode into the hippocampus. I was sitting in front of the oscilloscope, the instrument that displayed the electrical signals, and I also controlled the stimulators that could activate pathways into and out of the hippocampus. As I had done in Stanley Crain’s laboratory, I connected the recording electrode to a loudspeaker so that any electrical signal we might obtain could be heard as well as seen. We were trying to record from pyramidal cells, the major class of neurons in the hippocampus. These cells receive and process the information coming into the hippocampus and send it on to the next relay point. We also set up a camera to photograph the display screen of the oscilloscope.
Suddenly we heard the loud bang! bang! bang! of action potentials, a sound I recognized immediately from my experiments on crayfish. Alden had penetrated a cell! We quickly realized it was a pyramidal cell because the axons of these neurons are bundled into a pathway (called the fornix) that leads out of the hippocampus, and I had positioned electrodes on that pathway. Every stimulus I applied elicited a beautiful, large action potential. This method of stimulating the outgoing axon and causing pyramidal cells to fire proved to be a powerful way of identifying those cells. We also managed to excite pyramidal cells by stimulating the pathway that carries information
into
the hippocampus. We thus obtained a remarkable amount of information in the roughly ten minutes during which we recorded signals from pyramidal cells. We ran the camera continuously to ensure that every moment of the recording, every synaptic potential and every action potential in the pyramidal cells, was captured on film.
Alden and I were euphoric—we had obtained the first intracellular signals ever recorded from the region of the brain that stores our fondest memories! We almost danced around the lab. The mere accomplishment of recording from these cells successfully for several minutes met our most optimistic expectations. In addition, our data looked fascinating and somewhat different from what Eccles and Frank had found in the motor neurons of the spinal cord.
This experiment and the ones that followed were physically exhausting, sometimes lasting twenty-four hours. It was a good thing we had both just finished a medical internship, where working twenty-four hours at a stretch was not uncommon. We carried out three experiments a week and used the intervening two days, often only partial days because Jack was using the laboratory, to analyze data, to discuss the results, and just to talk. Many experiments were unsuccessful, but we eventually developed simple technical improvements that allowed us to obtain high-quality recordings once or twice a week.
By applying the powerful methodologies of cell biology to the hippocampus, Alden and I easily picked some low-hanging intellectual fruit. To begin with, we found that unlike motor neurons, a certain class of hippocampal neurons fires spontaneously, even without receiving instructions from sensory or other neurons. More interesting, we found that action potentials in the pyramidal cells of the hippocampus originate at more than one site within the cell. In the motor neuron, action potentials are initiated only at the base of the axon, where it emerges from the cell body. We had good evidence to suggest that action potentials in pyramidal cells of the hippocampus can also begin in the dendrites, and that they can be initiated in response to stimulation of the perforant pathway, an important direct synaptic input to the pyramidal cells from a part of the cortex called the entorrhinal cortex.
This proved to be an important discovery. Up to that time most neural scientists, including Dominick Purpura and Harry Grundfest, thought that dendrites could not be excited and therefore could not generate action potentials. Willifred Rall, a major theorist and model builder at NIH, had developed a mathematical model showing how the dendrites of motor neurons function. This model was based on the fundamental assumption that the cell membrane of dendrites is passive: it does not contain voltage-gated sodium channels and therefore cannot support an action potential. The intracellular signals we recorded were the first evidence to the contrary, and our finding later proved to be a general principle of neuronal function.
Our technical success and these intriguing findings brought enthusiastic encouragement and unstinting praise from our senior colleagues at NIH. John Eccles, who was emerging as the leading cellular physiologist in the mammalian brain, stopped by to see us when he visited NIH and was generous in his comments. Eccles invited Alden and me to join him in Australia to continue with him our work on the hippocampus, an offer we refused after much hesitation. Wade Marshall asked me to give a seminar at NIMH to summarize Alden’s and my efforts; I did, to a packed conference room, and it was warmly received. But even in our headiest moments, we realized that ours was a typical NIH story. Young, inexperienced people were given the opportunity to try things on their own, knowing that wherever they turned, experienced people were available to help.
Not everything was wine and roses, however. Soon after I arrived, another young scientist, Felix Strumwasser, went to work at a neighboring laboratory. Unlike the rest of the young research associates, who were medical doctors, Felix had a Ph.D. in neurophysiology from the University of California, Los Angeles. While most of us knew relatively little about brain science, Felix was extremely knowledgeable. We became good friends and had dinner at each other’s homes. I learned a great deal from him. In fact, in my conversations with Felix, I sharpened my thinking about how to tackle neurobiological studies of learning. Felix also got me to think about the hypothalamus, a region of the brain concerned with emotional expression and hormone secretions. The hypothalamus was becoming important in clinical discussions of how to treat stress and mental depression.