Junk DNA: A Journey Through the Dark Matter of the Genome (31 page)
Read Junk DNA: A Journey Through the Dark Matter of the Genome Online
Authors: Nessa Carey

i
This gene is called FTO or ‘fat mass and obesity associated’.
j
This gene is called IRX3, or Iroquois homeobox protein 3.
16. Lost in Untranslation
There are few crimes lower than deliberately hurting a child. In many countries, staff in emergency departments are trained to look for patterns of unexplained injuries including fractures in babies and toddlers. Often such a medical history will result in children being taken into care, little or no parental access, and ultimately prosecution and possibly imprisonment of one or both parents.
Protection of a child is of course paramount. But imagine the nightmare for parents if this happens to them and they are entirely innocent, because the fractures are due to an undetected medical condition.
1
Although the number of such miscarriages of justice is small compared with genuine cases of child abuse, the effects for the family are devastating. Loss of liberty, marital breakdown, social exclusion and, most heartbreakingly, the loss of parent–child contact.
A genetic condition can and has led to this misdiagnosis of child battery on more than one occasion. The disorder is called osteogenesis imperfecta, but it’s more commonly known as brittle bone disease.
2
Patients with brittle bone disease suffer fractures very easily, sometimes from mild traumas that might not even cause much of a bruise in a healthy child. The same bones may break repeatedly, and they may heal imperfectly, so that the affected person becomes increasingly disabled over time.
We might think that this condition should be very recognisable, making it rather strange that parents are sometimes wrongly accused of hurting their children. But there are a number of factors that complicate the picture. The first is that brittle bone disease
affects about six or seven children in 100,000. A doctor may simply have never encountered the condition, especially if they are relatively new to emergency medicine. But sadly, they probably will encounter child battery and so are more likely to have this as a default diagnosis.
The diagnosis is also complicated because there are at least eight different types of brittle bone disease, varying in their severity and the fine details of the presentation. At the most extreme, babies may suffer fractures even before they are born. The different forms of brittle bone disease are caused by mutations in different genes. The most common ones are defects in collagens, proteins that are important for making sure bones are flexible. Although we often think of bones as very rigid, it’s important that they have some flexibility, so that they bend rather than break in response to movement. It’s the same principle behind why we teach children not to climb on dead trees, because the inflexible, dried-out branches are more likely to break than the green, bendy limbs of living trees.
In most cases of brittle bone disease, only one copy of a gene is mutated. The other copy (because we inherit a copy from each parent) is fine. But having one normal copy isn’t enough to compensate for the effects of the ‘bad’ gene. Usually when this happens we expect to see a disorder in not just the child, but also in one parent. This is the parent who passes on the condition to their baby. But if the mutation is a new one, created during the production of eggs or sperm, a child can be affected without their parent having any symptoms. This tends to be particularly the case in the very severe forms of brittle bone disease. This makes it harder for doctors in an emergency room to recognise that they are looking at a condition caused by a mutation.
But if doctors do suspect that a baby may be suffering from brittle bone disease, they can order genetic tests to try to confirm their diagnosis. The genetic diagnosis will involve analysing the
sequences of the genes that are known to be mutated in brittle bone disease. Scientists will prioritise the order in which they sequence the genes by looking at the details of the patient’s symptoms, and deciding which form of brittle bone disease they think they have. Then they’ll sequence the most likely genes first, looking for mutations that alter the proteins required for strong healthy bones.
This usually works well. But inevitably we find that there are some patients with all the symptoms of brittle bone disease but who don’t have any mutations that alter the amino acid sequence in the proteins known to be involved in this condition. This is exactly the situation that faced scientists trying to understand the cause of a specific class of brittle bone disease
a
in a small number of Korean families. In this class of cases, there are characteristic patterns of fractures, but also a very strange after-effect. When the bones are damaged, either by a fracture itself or by medical intervention to repair a break, the patient’s body responds in an unusual way. It lays down too much calcium around the injury site, creating an obvious cloudy effect visible on an X-ray.
At the same time, other researchers were analysing a child from a German family, who had the same highly unusual type of brittle bone disease. Remarkably, the cases in both Korea and Germany were caused by exactly the same mutation. Just one base pair among the 3 billion the affected children inherited from each parent was altered. And the alteration that caused this disease was not in the amino acid-causing region of a gene. It was in junk DNA.
The beginning and the end
The mutation lay in a region of junk we have already encountered. In Chapter 2, we saw how protein-coding genes are composed
of modules. The modules are initially all copied into messenger RNA and various modules are joined together. Regions that don’t code for protein are removed during this ‘splicing’ process (
see page 16
).
But two regions of junk DNA always remain in the mature messenger RNA. These were shown in Figure 2.5 and Figure 16.1 depicts them again. Because these regions at the beginning and end of the messenger RNA are retained but never translated into protein, they are known as untranslated regions.
b
Although they don’t contribute to the amino acid sequence of the normal protein, researchers are identifying new ways in which these untranslated regions contribute to protein expression and to human health and disease.
The researchers in Korea analysed the DNA sequences of nineteen patients. Thirteen of these came from three affected families,
and the other six were single cases. Each of the nineteen patients had a change from a C base to a T base in the untranslated region at the start of the protein-coding region of a specific gene.
c
This change was just fourteen bases away from the start of the protein-coding region of the messenger RNA. They didn’t detect this C to T change in any of the unaffected family members or in 200 unrelated people from the same ethnic background.
3
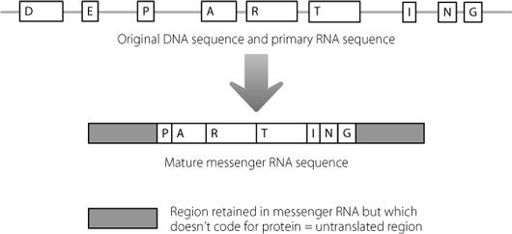
Figure 16.1
Even after the amino acid-coding regions of a messenger RNA have been spliced together, there is still some junk RNA which is retained in the molecule, at the beginning and end.
At about the same time, the researchers 5,000 miles away in Germany found exactly the same mutation in a young girl with the same type of brittle bone disease, and in another unrelated patient. In both cases, it was a fresh mutation. It wasn’t present in the parents and must have arisen during the production of eggs or sperm.
4
The scientists analysed the same region of the genome from over 5,000 unaffected people and found no one with this change.
There is a bit of a puzzle when we look at our image of messenger RNA in Figure 16.1. In the diagram the protein-coding regions and the untranslated regions have been drawn so that they look different from each other. But this isn’t what they are like in the cell. In reality, they look the same at the sequence level, because they are just formed from RNA bases.
For anyone fluent in written English, the following is pretty easy to decipher:
Iwanderedlonelyasacloud
Even though all the letters have been run together, we can recognise where individual words start and stop. The same is true for the cell, which is able to tell the difference between the sequences in the untranslated regions and in the amino acid-coding regions of a messenger RNA.
Translation of messenger RNA to create protein is carried out
at the ribosomes, in a process that we met in Chapter 11. The messenger RNA is fed through the ribosome, starting at the beginning of the messenger RNA molecule. Nothing much happens until the ribosome reads a particular three-base sequence, AUG (as mentioned in Chapter 2, the T base in DNA is always replaced by a slightly different base called U in RNA). This signals to the ribosome that it’s time to start joining up amino acids to create a protein.
Using our example from above, it would be as if we looked at a piece of text that read as:
dbfuwjrueahuwstqhwIwanderedlonelyasacloud
The capital I acts as the signal to us to start reading proper words, fulfilling a similar purpose to the AUG that signals the start of translation.
In the genes of the Korean and German patients with brittle bone disease, there is a point at which the normal DNA sequence in the untranslated region changes from ACG to ATG (which will be AUG in RNA). The consequence is that the ribosomes start the protein chain too early. This is shown in Figure 16.2.
This results in a strange phenomenon where junk RNA is changed to protein-coding RNA. This adds an extra five amino acids to the start of the normal protein, as shown in Figure 16.3. The protein involved in this type of brittle bone disease is one that has parts inside and outside the cell. The alteration in the junk DNA adds an extra five amino acids to a part of the protein that is outside the cell.
It’s not quite clear why these five amino acids cause the symptoms of the disease. Previous experiments in rodents had shown that too much or too little of this protein leads to defects in the skeleton, so it’s clear that having exactly the right amount of the protein is important.
5
The extra five amino acids are on a part of
the protein that we would expect might bind to other proteins or molecules that signal to the bone cells. It may be that having these extra five amino acids stops the mutant protein from responding properly, like putting chewing gum on the sensor of a smoke detector.
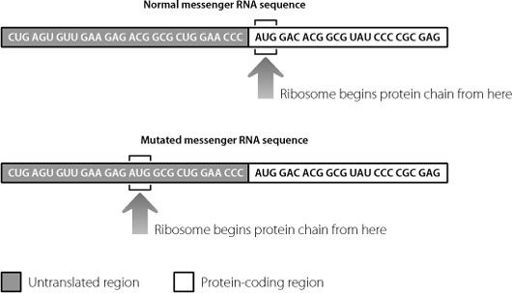
Figure 16.2
A mutation in the untranslated junk region at the beginning of the messenger RNA mis-directs the ribosome. The ribosome begins sticking amino acids together too early, creating a protein with an extraneous sequence at the beginning.
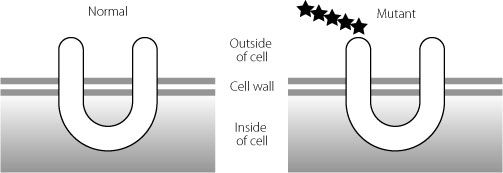
Figure 16.3
The U-shaped protein on the right has an extra five amino acids at the beginning, represented by stars. These extra amino acids probably influence which other molecules can interact with this protein.